关键词:重组自交系; 两系杂交粳稻; 杂种优势; 籼粳成分 Relationship between Indica-Japonica Index of Parents and Heterosis of Hybrid and Its Genetic Basis in Japonica Two Line Hybrid Rice YU Ya-Hui1,2, LIU Yu1, LI Zhen-Yu1, CHEN Guang-Hong1, XU Zheng-Jin2, TANG Liang2,*, MAO Ting1, XU Hai2 1 Liaoning Institute of Saline-Alkali Land Utilization, Panjin 124010, China
2 Rice Research Institute, Shenyang Agricultural University, Shenyang 110866, China
Fund:This study was supported by the Cultivation Plan for Youth Agricultural Science and Technology Innovative Talents of Liaoning Province (2015034, 2014046) and the Science and Technology Items of Liaoning Province (2014020004-101) AbstractIn order to further reveal the relationship between composition of subspecies lineage and heterosis in Oryza sativa L., we built a bridge for morphological index, Cheng’s index (Chi) and molecular index, japonica discrimination values ( Dj) based on two populations, a recombinant inbred lines (RILs), derived from the cross between indica cultivar Qishanzhan and japonica cultivar Akihikari, and japonica photo-thermo sensitive genic male sterile (PTGMS) line GB028S and their F1 hybrids. Two indexs achieved relatively consistent results in two populations independently. Significant curvilinear relationship appeared among Dj values, F1 yield and yield heterosis. The peaks of F1 yield and yield heterosis were detected in Dj interval from 0.55 to 0.70 and from 0.50 to 0.65 respectively, indicating that 0.55-0.65 is the preferential Dj interval for producing high yield and heterosis of F1 hybrids. Compositions of subspecies lineage on Chr.8, Chr.11, and Chr.12 were closely related with yield and heterosis of F1,however, the relationship among parent’s genetic distance, F1 yield and heterosis was not significant.
Keyword:RIL; Two-line hybrid rice; Heterosis; Indica-japonica composition Show Figures Show Figures
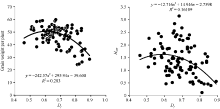
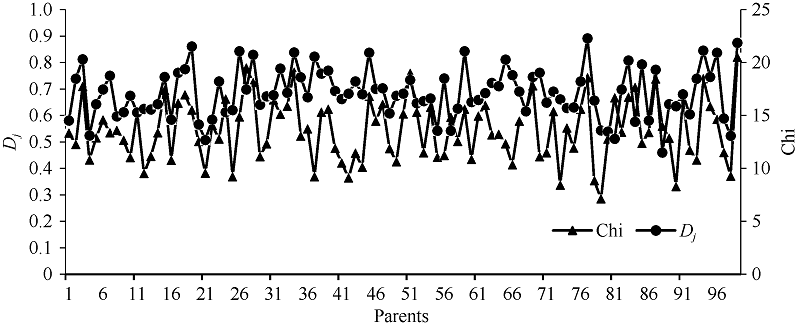
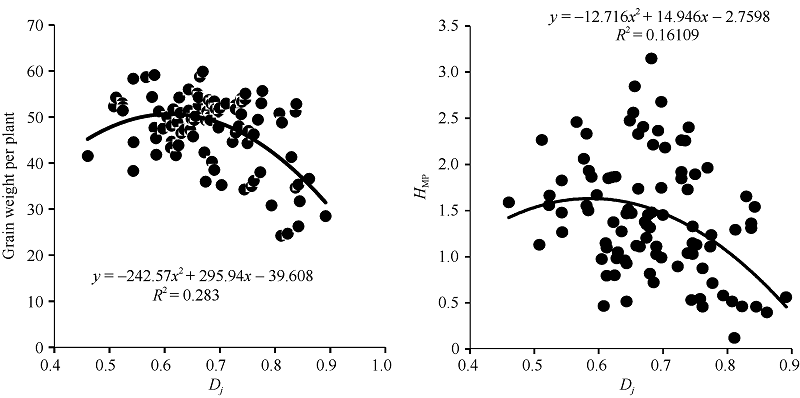
图2 RIL的Dj与F1单株产量和杂种优势的相关分析Dj: 偏粳系数; HMP: 中亲优势。Fig. 2 Relationship of RILs’ Dj with yield and HMP in F1Dj: Deviation japonica coefficient; HMP: Mid-parent heterosis.
表1 Table 1 表1(Table 1)
表1 RIL籼粳成分及亲本间遗传距离与F1产量性状和杂种优势的相关分析 Table 1 Relationship of RILs’ indica-japonica index and parental genetic distance with traits and heterosis in F1
性状 Trait
程氏指数 Cheng’ s index
偏粳系数 Dj
遗传距离 GD
X1
X2
X1
X2
X1
X2
单株产量 Grain weight per plant
-0.051
0.082
-0.287* *
-0.211*
-0.049
0.038
穗数 Panicles per plant
0.013
-0.309* *
-0.074
-0.164
-0.005
-0.071
穗粒数 Grains per panicle
-0.340* *
0.077
-0.261* *
-0.072
-0.130
-0.015
千粒重 1000-grain weight
0.302* *
0.014
0.165
-0.015
-0.051
-0.154
结实率 Seed setting rate
0.255*
-0.084
-0.003
-0.240*
0.084
-0.056
株高 Plant height
-0.152
0.041
-0.236*
-0.152
0.146
0.103
穗长 Panicle length
-0.086
0.009
-0.146
-0.080
-0.056
-0.079
着粒密度 Grain density
-0.276* *
-0.087
-0.194
-0.166
-0.117
-0.058
* , * * mean significant correlation at the 0.05 and 0.01, probability levels, respectively; X1: Relationship of parental indica-japonica index and GD with F1yield; X2: Relationship of parental indica-japonica index and GD with F1heterosis. * 和* * 表示在0.05、0.01水平上显著相关; X1: RIL籼粳成分及亲本间遗传距离与F1产量性状的相关系数; X2: RIL籼粳成分及亲本间遗传距离与F1产量性状杂种优势的相关系数。
表1 RIL籼粳成分及亲本间遗传距离与F1产量性状和杂种优势的相关分析 Table 1 Relationship of RILs’ indica-japonica index and parental genetic distance with traits and heterosis in F1
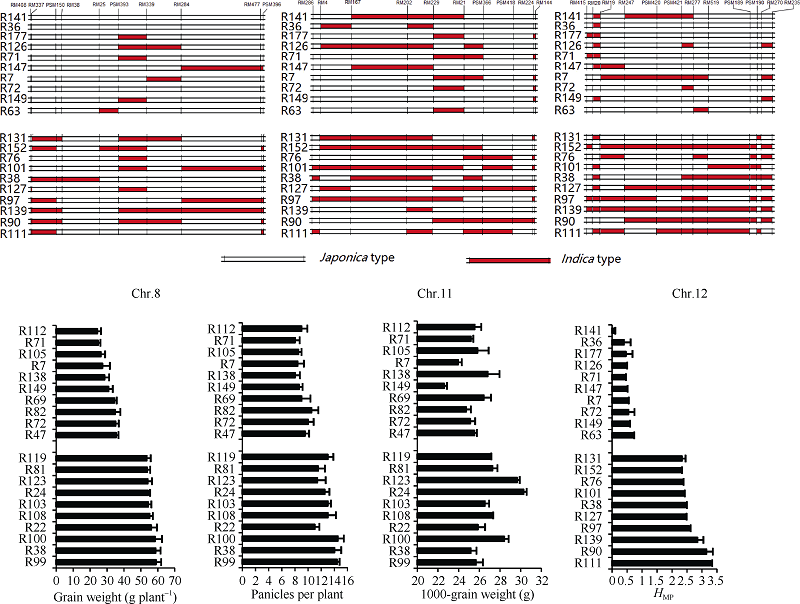
4 结论利用程氏指数法及分子标记法的偏粳系数在群体籼粳分类上具有较高的一致性。父本RIL的偏粳系数Dj值与单株产量及其杂种优势存在二次曲线关系。当父本的偏粳系数为0.55~0.65时, 杂种F1单株产量和杂种优势潜力较大。父本Chr.8、Chr.11和Chr.12上部分区段的籼粳成分与杂种F1产量及杂种优势关系密切。双亲的遗传距离与F1产量和相关性状及杂种优势没有明显的关系。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
徐海, 陶士博, 唐亮, 张文忠, 赵明辉, 徐凡. 栽培稻的籼粳分化与杂交育种研究进展. 沈阳农业大学学报, 2012, 43: 704-710XuH, Tao SB, TangL, Zhang WZ, Zhao MH, XuF. Research progress of differentiation and hybrid breeding between indica and japonica rices. J Shenyang Agric Univ, 2012, 43: 704-710 (in Chinese with English abstract)[本文引用:1]
[2]
姜健, 李金泉, 徐正进. 水稻籼粳杂交育种研究进展. 吉林农业科学, 2003, 28: 9-14JiangJ, Li JQ, Xu ZJ. The advancement of hybrid breeding between indica and japonica rices. J Jilin Agric Sci, 2003, 28: 9-14 (in Chinese with English abstract)[本文引用:2]
[3]
陈立云, 雷东阳, 唐文帮, 邓化冰, 肖应辉, 张桂莲. 论杂交水稻的育种方法. 湖南农业大学学报(自然科学版), 2015, 41: 1-4Chen LY, Lei DY, Tang WB, Deng HB, Xiao YH, Zhang GL. Review on the methods of hybrid rice breeding. J Hunan Agric Univ (Nat Sci), 2015, 41: 1-4 (in Chinese with English abstract)[本文引用:1]
[4]
陈立云, 雷东阳, 唐文帮, 邓化冰, 肖应辉, 张桂莲. 中国杂交水稻发展面临的挑战与策略. 杂交水稻, 2015, 30(5): 1-4Chen LY, Lei DY, Tang WB, Deng HB, Xiao YH, Zhang GL. Challenges and strategies of hybrid rice development in China. Hybrid Rice, 2015, 30(5): 1-4 (in Chinese with English abstract)[本文引用:1]
[5]
BotsteinD, White RL, SkolnickM, Davis RW. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am Soc Human Genet, 1980, 32: 314-331[本文引用:1]
[6]
毛艇, 徐海, 郭艳华, 朱春杰, 陈凯, 王嘉宇, 徐正进. 利用SSR分子标记进行水稻籼粳分类体系的初步构建. 华北农学报, 2009, 24: 119-124MaoT, XuH, Guo YH, Zhu CJ, ChenK, Wang JY, Xu ZJ. Establishment of subspecies classification of indica and japonica system by SSR markers. Acta Agric Boreali-Sin, 2009, 24: 119-124 (in Chinese with English abstract)[本文引用:1]
[7]
NeiM, Li WH. Mathematical model for studying genetic variation in terms of restriction endonuclease. Proc Nat Acad Sci USA, 1979, 76: 5269-5273[本文引用:1]
[8]
陈跃进, 张桂权, 卢永根. 利用微卫星分子标记法研究水稻亲缘关系. 湖南农业大学学报(自然科学版), 2007, 33: 258-261Chen YJ, Zhang GQ, Lu YG. Genetic relationship of rice based on STMS analysis. J Hunan Agric Univ (Nat Sci), 2007, 33: 258-261 (in Chinese with English abstract)[本文引用:1]
[9]
卢宝荣, 蔡星星, 金鑫. 籼稻和粳稻的高效分子鉴定方法及其在水稻育种和进化研究中的意义. 自然科学进展, 2009, 16: 628-638Lu BR, Cai XX, JinX. Efficient indica and japonica rice identification based on the InDel molecular method: its implication in rice breeding and evolutionary research. Prog Nat Sci, 2009, 19: 628-638 (in Chinese with English abstract)[本文引用:1]
[10]
刘丹, 王嘉宇, 柴永山, 孙玉友, 孙国宏, 冯章丽, 陈温福. 利用籼粳交RILs群体的水稻粒形QTLs定位. 沈阳农业大学学报, 2015, 46: 385-390LiuD, Wang JY, Cai YS, Sun YY, Sun GH, Feng ZL, Chen WF. Mapping of the QTLs for rice grain shape using recombinant inbred lines of indica-japonica hybridization in rice. J Shenyang Agric Univ, 2015, 46: 385-390 (in Chinese with English abstract)[本文引用:1]
[11]
SunJ, LiuD, Wang JY, Ma DR, TangL, GaoH, Xu ZJ, Chen WF. The contribution of intersubspecific hybridization to the breeding of super high-yielding japonica rice in northeast China. Theor Appl Genet, 2012, 125: 1149-1157[本文引用:1]
[12]
张培江, 才宏伟, 李焕朝, 杨联松, 张德泉, 白一松, 胡兴明, 许传万. RAPD分子标记水稻遗传距离及其与杂种优势的关系. 安徽农业科学, 2000, 28: 697-700Zhang PJ, Cai HW, Li HC, Yang LS, Zhang DQ, Bai YS, Hu XM, Xu CW. Genetic distance detected with RAPD markers among rice cultivars (Oryza sativa L. ) and its relation to heterosis. J Anhui Agric Sci, 2000, 28: 697-700 (in Chinese with English abstract)[本文引用:1]
[13]
王彦荣, 邱福林, 华泽田, 代贵金. 北方杂交粳稻亲本籼粳成分与杂种产量和稻米品质性状的关系. 中国水稻科学, 2009, 24: 145-150Wang YR, Qiu FL, Hua ZT, Dai JG. Relationship between indica-japonica index of parent s and hybrid yield and rice quality traits in japonica hybrid rice in northern China. Chin J Rice Sci, 2009, 24: 145-150 (in Chinese with English abstract)[本文引用:1]
[14]
Li YB, Fan CC, Xing YZ, Jiang YH, Luo LJ, SunL, ShaoD, Xu CJ, Li XH, Xiao JH, He YQ, Zhang QF. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet, 2011, 43: 1266-1269[本文引用:1]
[15]
王明军, 王云月, 陆春明, 杨慧, 王云涛. 利用籼粳稻特异InDel标记分析云南糯稻品种的籼粳特性. 云南农业大学学报, 2010, 25: 333-337Wang MJ, Wang YY, Lu CM, YangH, Wang YT. Characteristic of Yunnan glutinous rice varieties revealed by the indica-japonica specified insertion/deletion (InDel) molecular markers. J Yunnan Agric Univ, 2010, 25: 333-337 (in Chinese with English abstract)[本文引用:1]
[16]
赵小燕, 叶胜海, 李小华, 翟荣荣, 余鹏, 金庆生, 张小明. InDel标记鉴定水稻籼粳属性及预测杂种优势. 浙江农业学报, 2015, 27: 1309-1316Zhao XY, Ye SH, Li XH, Zhai RR, YuP, Jin QS, Zhang XM. Identification of indica-japonica attribute and prediction of heterosis using InDel markers in rice. Acta Agric Zhejiangensis, 2015, 27: 1309-1316 (in Chinese with English abstract)[本文引用:1]
[17]
徐群, 许红云, 魏兴华, 汤圣祥, 李东宣, 陈丽娟. 基于SSILP、InDel和SSR标记的杂草稻籼粳分类. 中国水稻科学, 2012, 26: 686-692XuQ, Xu HY, Wei XH, Tang SX, Li DX, Chen LJ. SSILP, InDel and SSR markers-based indica-japonica classification for weedy. Chin J Rice Sci, 2012, 26: 686-692 (in Chinese with English abstract)[本文引用:1]
[18]
Lou XJ, Fu YC, Zhang PJ, WuS, TianF, Liu JY, Zhu ZF, Yang JS, Sun CQ. Additive and over-dominant effects resulting from epistatic loci are the primary genetic basis of heterosis in rice. J Integr Plant Biol, 2009, 51: 393-408[本文引用:1]
[19]
陈深广, 沈希宏, 曹立勇, 占小登, 冯跃, 吴伟明, 程式华. 水稻产量性状杂种优势的QTL定位. 中国农业科学, 2010, 43: 4983-4990Chen SG, Shen XH, Cao LY, Zhan XD, FengY, Wu WM, Cheng SH. QTL mapping for heterosis of yield traits in rice. Sci Agric Sin, 2010, 43: 4983-4990 (in Chinese with English abstract)[本文引用:1]
[20]
赵秀琴, 朱苓华, 徐建龙, 黎志康. 灌溉与自然降雨条件下水稻高代回交导入系产量QTL的定位. 作物学报, 2007, 33: 1536-1542Zhao XQ, Zhu LH, Xu JL, Li ZK. QTL mapping of yield under irrigation and rainfed field conditions for advanced backcrossing intrigression lines in rice. Sci Agric Sin, 2007, 33: 1536-1542 (in Chinese with English abstract)[本文引用:2]
[21]
陈冰嬬, 石英尧, 崔金腾, 钱益亮, 刘海燕, 张力科, 王辉, 高用明, 朱苓华, 黎志康. 利用BC2F2 高代回交群体定位水稻籽粒大小和形状QTL. 作物学报, 2008, 34: 1299-1307Chen BX, Shi YY, Cui JT, Liu HY, Zhang LK, WangH, Gao YM, Zhu LH, Li ZK. QTL detection of grain size and shape with BC2F2 advanced backcross population of rice (Oryza sativa L. ). Sci Agric Sin, 2008, 34: 1299-1307 (in Chinese with English abstract)[本文引用:1]
[22]
Li DJ, Sun CQ, Fu YC, LiC, Zhu ZF, ChenL, Cai HW, Wang XK. Identification and mapping of genes for improving yield from Chinese common wild rice (O. rufipogon Griff. ) using advanced backcross QTL analysis. Chin Sci Bull, 2002, 47: 1533-1537[本文引用:1]
[23]
ChaiL, ZhangJ, Pan XB, ZhangF, Zheng TQ, Zhao XQ, Wang WS, JauharA, Xu JL, Li ZK. Advanced backcross QTL analysis for the whole plant growth duration salt tolerance in rice (Oryza sativa L. ). J Integr Agric, 2014, 13: 1609-1620[本文引用:1]
[24]
赵芳明, 张桂权, 曾瑞珍, 杨正林, 凌英华, 桑贤春, 何光华. 基于单片段代换系的水稻粒型QTL加性及上位性效应分析. 作物学报, 2011, 37: 469-476Zhao FM, Zhang GQ, Zeng RL, Yang ZL, Ling YH, Sang XC, He GH. Analysis of epistatic and additive effects of QTLs for grain shape using single segment substitution lines in rice (Oryza sativa L. ). Acta Agron Sin, 2011, 37: 469-476 (in Chinese with English abstract)[本文引用:1]
[25]
王智权, 江玲, 尹长斌, 王晓玲, 雷建国, 肖宇龙, 刘喜, 刘世家, 陈亮明, 余传元, 万建民. 水稻产量相关农艺性状杂种优势位点的定位. 中国水稻科学, 2013, 27: 569-576Wang ZQ, JiangL, Yin CB, Wang XL, Lei JG, Xiao YL, LiuX, Liu SJ, Chen ML, Yu CY, Wan JM. QTL mapping of heterotic loci of yield-related traits in rice. Chin J Rice Sci, 2013, 27: 569-576 (in Chinese with English abstract)[本文引用:2]
[26]
徐建龙, 薛庆中, 罗利军, 黎志康. 水稻单株有效穗数和每穗粒数的QTL剖析. 遗传学报, 2001, 28: 752-759Xu JL, Xue QZ, Luo LJ, Li ZK. QTL dissection of panicle number per plant and spikelet number per panicle in rice (Oryza sative L. ). Acta Genet Sin, 2001, 28: 752-759 (in Chinese with English abstract)[本文引用:1]
[27]
辛业芸, 袁隆平. 超级杂交稻两优培九产量杂种优势标记与QTL分析. 中国农业科学, 2014, 47: 2699-2714Xin YY, Yuan LP, Heterosis loci and QTL of super rice Liangyoupeijiu yield by using molecular marker. Sci Agric Sin, 2014, 47: 2699-2714 (in Chinese with English abstract)[本文引用:1]
[28]
穆平, 黄超, 李君霞, 刘立峰, 刘弋菊, 李自超. 低磷胁迫下水稻产量性状变化及其QTL定位. 作物学报, 2008, 34: 1137-1142MuP, HuangC, Li JX, Liu LF, Liu YJ, Li ZC. Yield trait variation and QTL mapping in a DH population of rice under phosphorus deficiency. Acta Agron Sin, 2008, 34: 1137-1142 (in Chinese with English abstract)[本文引用:1]
[29]
李任华, 徐才国, 何予卿, 袁隆平, 王象坤. 水稻亲本遗传分化程度与籼粳杂种优势的关系. 作物学报, 1998, 24: 564-576Li RH, Xu CG, He YQ, Yuan LP, Wang XK. Relationship between the extent of genetic differentiation in parental lines and heterosis of indica-japonica hybrid rice. Acta Agron Sin, 1998, 24: 564-576 (in Chinese with English abstract)[本文引用:1]
[30]
廖伏明, 周坤炉, 阳和华, 徐秋生. 杂交水稻亲本遗传差异及其与杂种优势关系. 中国水稻科学, 1998, 12: 193-199Liao FM, Zhou KL, Yang HH, Xu QS. Genetic difference of parents and its relation to heterosis in hybrid rice. Chin J Rice Sci, 1998, 12: 193-199 (in Chinese with English abstract)[本文引用:1]
[31]
孙传清, 姜廷波, 陈亮, 吴长明, 李自超, 王象坤. 水稻杂种优势与遗传分化关系的研究. 作物学报, 2000, 26: 641-649Sun CQ, Jiang TB, ChenL, Wu CM, Li ZC, Wang XK. Studies on the relationship between heterosis and genetic differentiation in hybrid rice (Oryza sativa L. ). Acta Agron Sin, 2000, 26: 641-649 (in Chinese with English abstract)[本文引用:1]
[32]
朱作峰, 孙传清, 姜廷波, 付强, 王象坤. 水稻品种SSR、RFLP及其与杂种优势的关系比较研究. 遗传学报, 2001, 28: 738-745Zhu ZF, Sun CQ, Jiang TB, FuQ, Wang XK. The comparison of genetic divergences and its relationships to heterosis revealed by SSR and RFLP markers in rice (Oryza sativa L. ). Acta Genet Sin, 2001, 28: 738-745 (in Chinese with English abstract)[本文引用:1]
[33]
张涛, 韩磊, 徐建第, 蒋开锋, 吴先军, 汪旭东, 郑家奎. 杂交香稻亲本遗传距离与产量杂种优势的相关性研究. 中国农业科学, 2006, 39: 831-835ZhangT, HanL, Xu JD, Jiang KF, Wu XJ, Wang XD, Zheng JK. Correlation between genetic distance and yield heterosis of hybrid aromatic rice. Sci Agric Sin, 2006, 39: 831-835 (in Chinese with English abstract)[本文引用:1]
[34]
蔡健, 兰伟. AFLP标记与水稻杂种产量及产量杂种优势的预测. 中国农学通报, 2005, 21(4): 39-43CaiJ, LanW. Using of AFLP marker to predict the hybrid yield and yield heterosis in rice. Chin Agric Sci Bull, 2005, 21(4): 39-43 (in Chinese with English abstract)[本文引用:1]
[35]
赵庆勇, 朱镇, 张亚东, 赵凌, 陈涛, 张巧凤, 王才林. SSR标记遗传距离与粳稻杂种优势的相关性分析. 中国水稻科学, 2009, 23: 141-147Zhao QY, ZhuZ, Zhang YD, ZhaoL, ChenT, Zhang QF, Wang CL. Analysis on correlation between heterosis and genetic distance based on simple sequence repeat markers in japonica rice. Chin J Rice Sci, 2009, 23: 141-147 (in Chinese with English abstract)[本文引用:1]
[36]
王林友, 张礼霞, 勾晓霞, 范宏环, 金庆生, 王建军. 利用InDel标记鉴定浙优系列杂交稻籼粳属性和预测杂种优势. 中国农业科学, 2014, 47: 1243-1255Wang YL, Zhang LX, Gou XX, Fan HH, Jin QS, Wang JJ. Identification of indica-japonica attribute and prediction of heterosis of Zheyou hybrids rice using InDel molecular markers. Sci Agric Sin, 2014, 47: 1243-1255 (in Chinese with English abstract)[本文引用:1]
[37]
罗小金, 贺浩华, 彭小松, 余秋英, 孙俊立, 张洪亮, 李自超. 利用SSR标记分析水稻亲本间遗传距离与杂种优势的关系. 植物遗传资源学报, 2006, 7: 209-214Luo XJ, He HH, Peng XS, Yu QY, Sun JL, Zhang HL, Li ZC. Analysis on correlation between genetic distance of rice parents and heterosis with SSR molecular marker. J Plant Genet Resour, 2006, 7: 209-214 (in Chinese with English abstract)[本文引用:1]
[38]
游书梅, 曹应江, 郑家奎, 姜开峰, 张涛, 杨莉, 杨乾华, 万先齐, 罗婧. 杂交籼稻子粒QTL标记遗传距离与粒重、粒重杂种优势的相关性. 植物遗传资源学报, 2012, 13: 1050-1054You SM, Cao YJ, Zheng JK, Jiang KF, ZhangT, YangL, Wan XQ, LuoJ. Correlation between genetic distance based on grain weight QTL markers, grain weight and grain weight heterosis in indica hybrid rice. J Plant Genet Resour, 2012, 13: 1050-1054 (in Chinese with English abstract)[本文引用:1]
[39]
Melchinger AE, LeeM, Lamkey KR. Genetic diversity for restriction fragment length polymorphisms: Relation to estimated genetic effects in maize inbreds. Crop Sci, 1990, 30: 1033-1040[本文引用:1]
[40]
ZhouG, ChenY, YaoW, Zhang CJ, Xie WB, Hua JP, Xing YZ, Xiao JH, Zhang QF. Genetic composition of yield heterosis in an elite rice hybrid. Proc Natl Acad Sci USA, 2012, 109: 15847-15852[本文引用:1]
[41]
GuoY, Hong DL. Novel pleiotropic loci controlling panicle architecture across environments in japonica rice (Oryza sativa L. ). J Genet Genomics, 2010, 37: 531-542[本文引用:1]
[42]
何光华, 侯磊, 李德谋, 罗小英, 牛国清, 唐梅, 裴炎. 利用分子标记预测杂交水稻产量及其构成因素. 遗传学报, 2002, 29: 438-444He GH, HouL, Li DM, Luo XY, Niu GQ, TangM, PeiY. Prediction of yield and yield components in hybrid rice by using molecular markers. Acta Genet Sin, 2002, 29: 438-444 (in Chinese with English abstract)[本文引用:1]
[43]
Saghai MM, YangG. Correlation between molecular marker distance and hybrid performance in US southern long grain rice. Crop Sci, 1997, 37: 145-150[本文引用:1]
[44]
Zhao MF, Li XH, Yang JB. Relationship between molecular marker heterozygosity and hybrid performance in intra- and inter-subspecific crosses of rice. Plant Breed, 1999, 118: 139-144[本文引用:1]
[45]
Zha RM, Ling YH, Yang ZL, Zhao FM, Zhong BQ, XieR, Sang XC, He GH. Prediction of hybrid grain yield performances in indica rice (Oryza sativa L. ) with effect-increasing loci. Mol Breed, 2008, 22: 467-476[本文引用:1]
[46]
张小祥, 李爱宏, 肖宁, 潘存红, 余玲, 李育红, 黄年生, 刘广青, 周长海, 季红娟, 刘晓静, 王宝和, 赵步洪, 谭长乐, 戴正元. 运用籼粳特异分子标记划分杂交稻亲本群及杂种优势模式初探. 扬州大学学报(农业与生命科学版), 2014, 35(1): 55-60Zhang XY, Li AH, XiaoN, Pan CH, YuL, Li YH, Huang NS, Liu GQ, Zhou CH, Ji HJ, Liu XJ, Wang BH, Zhao BH, Tan CL, Dai ZY. Division of the hybrid rice parent group using the indica-japonica specific markers to study the pattern of heterosis. J Yangzhou Univ (Agric & Life Sci Edn), 2014, 35(1): 55-60 (in Chinese with English abstract)[本文引用:1]
[47]
XiaoJ, LiL. Genetic diversity and its relationship to hybrid performance and heterosis in rice as revealed by PCR-based markers. Theor Appl Genet, 1996, 92: 637-643[本文引用:1]
[48]
梁奎, 黄殿成, 赵凯铭, 阮方松, 谢辉, 马文霞, 洪德林. 杂交粳稻亲本产量性状优异配合力的标记基因型筛选. 作物学报, 2010, 36: 1270-1279LiangK, Huang DC, Zhao KM, Ruan FS, XieH, Ma WX, Hong DL. Screening marker genotypes with elite combining ability for yield traits in parents of hybrid japonica rice (Oryza sativa L. ). Acta Agron Sin, 2010, 36: 1270-1279 (in Chinese with English abstract)[本文引用:1]
[49]
黄殿成, 阮方松, 刘健, 谢辉, 赵凯铭, 梁奎, 江建华, 蔡勇林, 洪德林. 杂交粳稻亲本穗长和枝梗数性状优异配合力的标记基因型筛选. 南京农业大学学报, 2011, 34(2): 1-7Huang DC, Ruan FS, LiuJ, XieH, Zhao KM, LiangK, Jiang JH, Cai YL, Hong DL. Screening marker genotypes with elite combining ability for panicle length and branch numbers in parents of hybrid japonica rice (Oryza sativa L. ). J Nanjing Agric Univ, 2011, 34(2): 1-7 (in Chinese with English abstract)[本文引用:1]
[50]
江建华, 刘强明, 卢超, 张红, 刘晓丽, 党小景, 牛付安, Caleb BM, 赵凯铭, 洪德林. 粳稻杂种优势遗传基础剖析. 作物学报, 2012, 38: 2147-2161Jiang JH, Liu QM, LuC, ZhangH, Liu XL, Dang XJ, Niu FA, Caleb BM, Zhao KM, Hong DL. Genetic basis dissection of heterosis in japonica rice (Oryza sativa L. ). Acta Agron Sin, 2012, 38: 2147-2161 (in Chinese with English abstract)[本文引用:1]
[51]
张红, 江建华, 刘晓丽, 吉家曾, 牛付安, 张有富, 洪德林. 粳稻8个异交相关性状及其中亲优势的QTL定位与遗传分析. 中国水稻科学, 2013, 27: 247-258ZhangH, Jiang JH, Liu XL, Ji JZ, Niu FA, Zhang YF, Hong DL. QTL mapping and genetic analysis of eight outcrossing-related traits and its mid-parental-heterosis in japonica rice. Chin J Rice Sci, 2013, 27: 247-258 (in Chinese with English abstract)[本文引用:1]
[52]
Huang XH, Yang SH, Gong JY, ZhaoY, FengQ, GongH, Li WJ, QiL, Zhan QL, Cheng BY, Chen BY, Xia JH, ChenN, ZhongN, Hao ZN, Liu KY, Zhu CR, Zhu CR, HuangT, ZhaoQ, ZhangL, Fan DL, Zhou CC, Lu YQ, Wen QJ, Wang ZX, Li JY, HanB. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nat Commun, 2015, DOI: DOI:10.1038/ncomms7258[本文引用:1]
[53]
赵彦宏, 朱军, 徐海明, 杨剑, 高用明, 宋佑胜, 石春海, 邢永忠. 基于QTL定位的水稻有效穗数杂种优势预测. 中国水稻科学, 2007, 21: 350-354Zhao YH, ZhuJ, Xu MH, YangJ, Gao YM, Song YS, Shi CH, Xing YZ. Predicting heterosis of effective panicle number per plant based on QTL mapping in rice. Chin J Rice Sci, 2007, 21: 350-354 (in Chinese with English abstract)[本文引用:1]
 , 毛艇
, 毛艇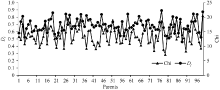

{kind=link}
{kind=link}
{kind=link}