关键词:高丹草; 叶片; 杂种优势; 蛋白质组 Analysis of Heterosis in Sorghum-Sudangrass Hybrid Seedlings Based on Proteomics HAN Ping-An1, LU Xiao-Ping1,*, MI Fu-Gui2, ZHANG Rui-Xia3, LI Mei-Na3, XUE Chun-Lei1, DONG Jing1, CONG Meng-Lu1 1 College of Agronomy, Inner Mongolia Agricultural University, Hohhot 010019, China
2 College of Ecology and Environmental Science, Inner Mongolia Agricultural University, Hohhot 010019, China
3 Hohhot Seed Management Station, Hohhot 010020, China
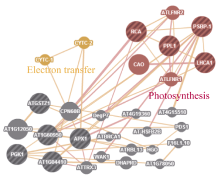
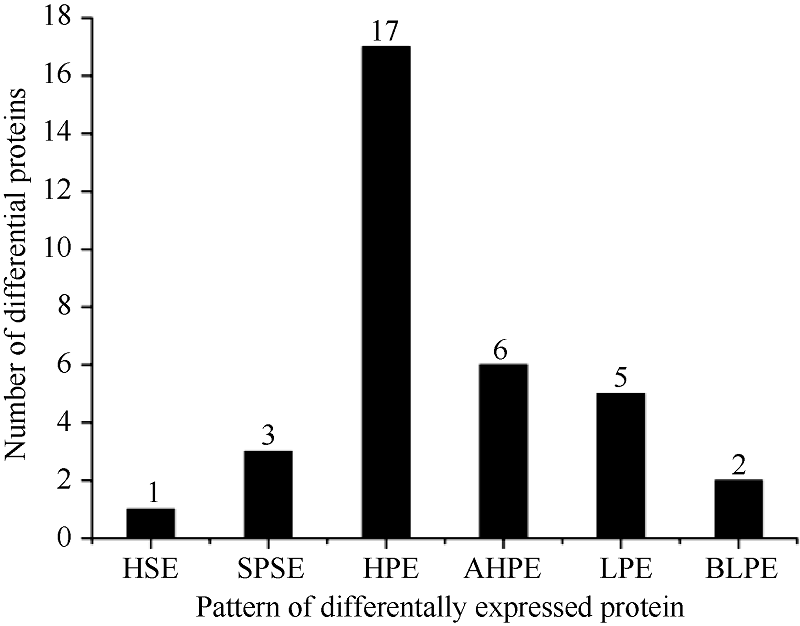
Fund:This study was supported by the National Natural Science Foundation of China (31160302, 31460375) and the Science and Technology Plan Projects of Hohhot (2012-major plans-8-2) AbstractSorghum-sudangrass hybrids are typically used for studying heterosis in forage crops. In this study, we carried out proteomic research on sorghum-sudangrass hybrids and their parents at the three-leaf stage by two dimensional electrophoresis-based proteomics and bioinformatic methods. More than 400 protein spots were detected, in which 34 proteins showed significant differences between hybrid and parents in expression, including dominant expression (showing three single-parent silent, seventeen high-parent and five low-parent expression) and overdominant expression (showing one hybrid-specific, six above-high-parent, two below-low-parent expression). Thus, we speculated that dominant and overdominant effects play key roles, and dominant effect is a major factor in the formation of heterosis in sorghum-sudangrass hybrid. Moreover, 27 out of 34 proteins were related to eight functional categories, i.e., photosynthesis, carbohydrate metabolism, stress response, ATP synthesis, protein synthesis, electron transfer, signal transduction and unknown. The up-regulated photosynthetic proteins were the biggest category, which indicates that photosynthesis in the leaves of sorghum-sudangrass hybrid is enhanced resultig in producing more organic matter, so that showing heterosis. The identified key node proteins in the interaction networks were the potential target proteins for future genetic manipulation of the specific proteins of heterosis. Our findings provide a theoretical basis on heterosis analysis of sorghum-sudangrass hybrids, which is potentially useful for other forage plants.
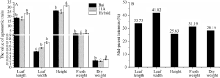
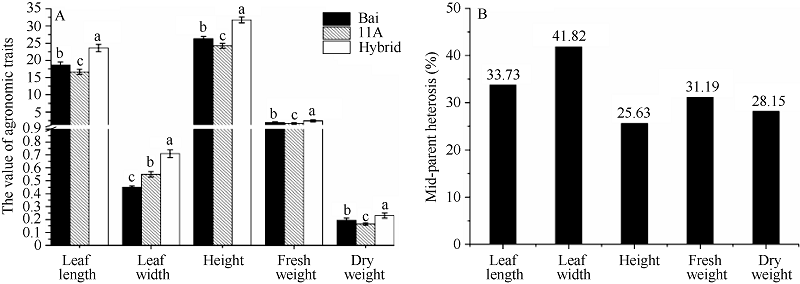
图2 高丹草及其亲本苗期杂种优势分析A: 高丹草与亲本各项指标的方差分析; B: 高丹草与亲本各项指标的中亲优势值。Fig. 2 Heterosis at seeding stage of sorghum-sudangrass hybrid and its parentsA: variance analysis of agronomic traits of sorghum-sudangrass hybrid and parents: B: mid-parent heterosis of sorghum-sudangrass hybrid and parents. Bars superscripted by different letters are significantly different at P < 0.05.
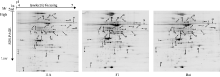
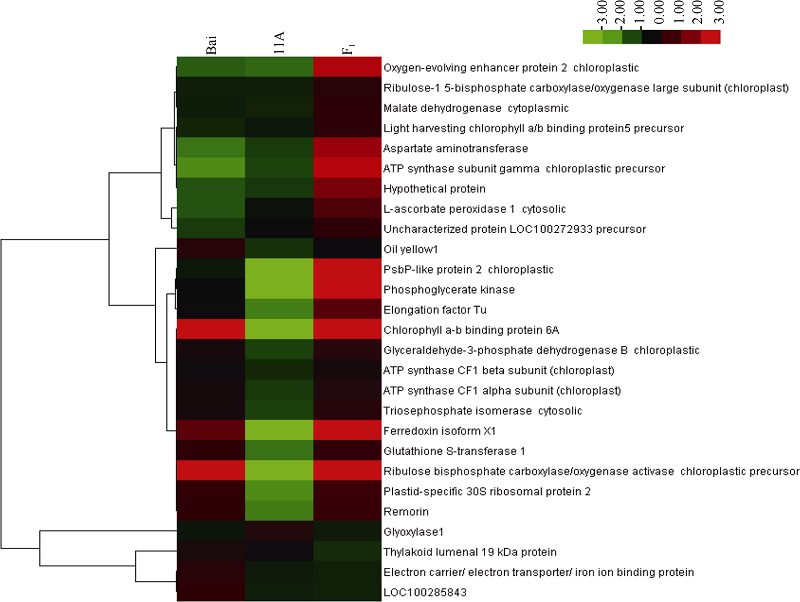
图6 差异蛋白聚类分析3列分别代表3种基因型( Bai, 11A, F1), 行代表单个蛋白, 左侧是对蛋白质聚类, 右侧是蛋白名称, 红色代表上调表达, 绿色代表下调蛋白。右边彩条的颜色强度代表不同蛋白表达量的变化Fig. 6 Clustering analysis of differentially expressed proteinsThe columns represent three genotypes (Bai, 11A, F1), the rows represent the individual protein. The protein cluster is on the left side. The name of protein is on the right side, the up- or down-regulated proteins are indicated in red and green, respectively. The intensity of the colors increases with increasing expression level as noted on the colorbar on the right side.
韩平安, 逯晓萍, 王亚男, 米福贵, 张瑞霞, 董婧. 基于重组自交系群体的高丹草抗倒高产种质的筛选. 中国草地学报, 2014, 36(5): 51-57Han PA, Lu XP, Wang YN, Mi HG, Zhang RX, DongJ. Screening of the lodging-resistance and high-yielding germplasm of sorghum sudan grass hybrid based on recombinant inbred line populations. Chin J Grassland , 2014, 36(5): 51-57 (in Chinese with English abstract)[本文引用:1]
[2]
Lu XP, Yun JF, Gao CP, SuryaA. Quantitative trait loci analysis of economically important traits in Sorghum bicolor S. sudanense hybrid. Can J Plant Sci, 2011, 91: 81-90[本文引用:1]
[3]
Bruce AB. The Mendelian theory of heredity and the augmentation of vigor. Science, 1910, 32: 627-628[本文引用:1]
[4]
DavenportC. Degeneration, albinism and inbreeding. Science, 1908, 28: 454-456[本文引用:1]
[5]
Crow JF. Alternative hypotheses of hybrid vigor. Genetics, 1948, 33: 477-487[本文引用:1]
[6]
Hull FH. Recurrent selection for specific combining ability in corn. Agron J, 1945, 37: 134-145[本文引用:1]
[7]
PowersL. An expansion of Jones’s theory for the explanation of heterosis. Am Nat, 1944, 78: 275-280[本文引用:1]
[8]
WilliamsW. Heterosis and the genetics of complex characters. Nature, 1959, 184: 527-530[本文引用:1]
[9]
Stupar RM, Springer NM. cis-transcriptional variation in maize inbred lines B73 and Mo17 leads to additive expression patterns in the F1 hybrid. Genetics, 2006, 173: 2199-2210[本文引用:1]
[10]
ZhangL, PengY, WeiX, DaiY, YuanD, LuY, PanY, ZhuZ. Small RNAs as important regulators for the hybrid vigour of super-hybrid rice. J Exp Bot, 2014, 65: 5989-6002[本文引用:1]
[11]
WangH, FengQ, ZhangM, YangC, ShaW, LiuB. Alteration of DNA methylation level and pattern in sorghum (Sorghum bicolor L. ) pure-lines and inter-line F1 hybrids following low-dose laser irradiation. J Photochem Photobiol B, 2010, 99: 150-153[本文引用:1]
[12]
GaoM, HuangQ, ChuY, DingC, ZhangB, SuX. Analysis of the leaf methylomes of parents and their hybrids provides new insight into hybrid vigor in populus deltoides. BMC Genet, 2014, 15(suppl-1): S8[本文引用:1]
[13]
SongX, NiZ, YaoY, XieC, LiZ, WuH, ZhangY, SunQ. Wheat (Triticum aestivum L. ) root proteome and differentially expressed root proteins between hybrid and parents. Proteomics, 2007, 7: 3538-3557[本文引用:1]
[14]
SongX, NiZ, YaoY, ZhangY, SunQ. Identification of differentially expressed proteins between hybrid and parents in wheat (Triticum aestivum L. ) seedling leaves. Theor Appl Genet, 2009, 118: 213-225[本文引用:1]
[15]
WangW, MengB, GeX, SongS, YangY, YuX, WangL, HuS, LiuS, YuJ. Proteomic profiling of rice embryos from a hybrid rice cultivar and its parental lines. Proteomics, 2008, 8: 4808-4821[本文引用:1]
[16]
进茜宁, 付志远, 丁冬, 刘宗华, 李卫华, 汤继华. 玉米杂交种先玉335 及其亲本种子萌发过程中胚芽蛋白质组学分析. 作物学报, 2011, 37: 1689-1694Jin XN, Fu ZY, DingD, Liu ZH, Li WH, Tang JH. Proteomic analysis of plumule in seed germination for an elite hybrid pioneer 335 and its parental lines in maize. Acta Agron Sin, 2011, 37: 1689-1694 (in Chinese with English abstract)[本文引用:1]
[17]
郭宝健, 隋志鹏, 李洋洋, 冯万军, 闫文文, 李慧敏, 孙其信, 倪中福. 玉米杂交种与亲本苗期叶片差异表达蛋白谱分析. 中国农业科学, 2013, 46: 3046-3054Guo BJ, Sui ZP, Li YY, Feng WJ, Yan WW, Li HM, Sun QX, Ni ZF. Differentially expressed protein profile of maize seedling leaves between hybrid and its parental lines. Sci Agric Sin, 2013, 46: 3046-3054 (in Chinese with English abstract)[本文引用:3]
[18]
郭宝健, 宋方威, 冯万军, 隋志鹏, 孙其信, 倪中福. 玉米杂交种与亲本雌穗花器官形成期蛋白差异表达谱分析. 作物学报, 2013, 39: 845-854Guo BJ, Song FW, Feng WJ, Sui ZP, Sun QX, Ni ZF. Differentially expressed protein profiling during ear floral development between maize hybrid and its parents. Acta Agron Sin, 2013, 39: 845-854 (in Chinese with English abstract)[本文引用:1]
[19]
Ajit KS, Syed IA, NabonitaS, RenuS. Differential proteomic analysis of salt stress response in sorghum bicolor leaves. J Environ Exp Bot, 2011, 71: 321-328[本文引用:1]
[20]
HanB, LiC, ZhangL, FangY, FengM, LiJ. Novel royal jelly proteins identified by gel-based and gel-free proteomics. J Agric Food Chem, 2011, 59: 10346-10355[本文引用:1]
[21]
HoeckerN, LamkemeyerT, SarholzB, PascholdA, FladererC, MadlungJ, WursterK, StahlM, Piepho HP, NordheimA, HochholdingerF. Analysis of non additive protein accumulation in young primary roots of a maize (Zea mays L. ) F1-hybrid compared to its parental inbred lines. Proteomics, 2008, 8: 3882-3894[本文引用:1]
[22]
ZhangC, YinY, ZhangA, LuQ, WenX, ZhuZ, ZhangL, LuC. Comparative proteomic study reveals dynamic proteome changes between superhybrid rice LYP9 and its parents at different developmental stages. J Plant Physiol, 2012, 16: 387-398[本文引用:1]
[23]
XiaY, NingZ, BaiG, LiR, YanG, Siddique KH, BaumM, GuoP. Allelic variations of a light harvesting chlorophyll a/b-binding protein gene (Lhcb1) associated with agronomic traits in barley. PLoS One, 2012, 7: e37573[本文引用:1]
[24]
Green BR, PicherskyE, KloppstechK. Chlorophyll a/b-binding proteins: an extended family. Trends Biochem Sci, 1999, 16: 181-186[本文引用:1]
[25]
HallM, KieselbachT, Sauer UH, Schröder WP. Purification, crystallization and preliminary X-ray analysis of PPD6, a PsbP-domain protein from Arabidopsis thaliana. Acta Crystallogr Sect F Struct Biol Cryst Commun, 2012, 68: 278-280[本文引用:1]
CsiszárJ, HorváthE, VáryZ, GalléÁ, BelaK, BrunnerS, TariI. Glutathione transferase supergene family in tomato: salt stress-regulated expression of representative genes from distinct GST classes in plants primed with salicylic acid. Plant Physiol Biochem, 2014, 78: 15-26[本文引用:1]
[32]
KumarS, Asif MH, ChakrabartyD, Tripathia RD, Dubeyb RS, TrivediP K. Expression of a rice Lambda class of glutathione S-transferase OsGSTL2, in Arabidopsis provides tolerance to heavy metal and other abiotic stress. J Hazard Mater, 2013, 248/249: 228-237[本文引用:1]
[33]
MoustakaJ, TanouG, Adamakis ID, Eleftheriou EP, Eleftheriou MM, MoustakasM. Leaf age-dependent photoprotective and antioxidative response mechanisms to paraquat-induced oxidative stress in Arabidopsis thaliana. Int J Mol Sci, 2015, 16: 13989-14006[本文引用:1]
[34]
McCartyR E. A plant biochemist's view of H+-ATPases and ATP syntheses. J Exp Bot, 1992, 172: 431-441[本文引用:1]
[35]
vonBallmoos C, DimrothP. Two distinct proton binding sites in the ATP synthase family. Biochemistry, 2007, 46: 11800-11809[本文引用:1]
[36]
TaizL, Zeiger E. Plant Physiology. Sinauer Associates Inc. , USA, 2002. pp 145-150[本文引用:1]
[37]
ZhangH, LuoM, Day RC, Talbot MJ, IvanovaA, Ashton AR, Chaudhury AM, Macknight RC, HrmovaM, Koltunow AM. Developmentally regulated HEART STOPPER, a mitochondrially targeted L18 ribosomal protein gene, is required for cell division, differentiation, and seed development in Arabidopsis. J Exp Bot, 2015, 66: 5867-5880[本文引用:1]
 , 米福贵
, 米福贵








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}