关键词:杂交粳稻; 骨干亲本; 产量性状配合力; 标记位点基因型 Identifying SSR Marker Locus Genotypes with Elite Combining Ability for Yield Traits in Backbone Parents of JaponicaHybrid Rice ( Oryza sativaL.) in Jianghuai Area XIE Hui1,2, DANG Xiao-Jing1, LIU Er-Bao1, ZENG Si-Yuan1, HONG De-Lin1,* 1State Key Laboratory of Crop Genetics and Germplasm Enhancement, Nanjing Agricultural University, Nanjing 210095, China
2 China National Japonica Rice Research and Development Center, Tianjin 300457, China
Fund:This study was supported by China National High-tech Research and Development Program (2010AA101301), the Doctoral Found of Educational Ministry (20110097110038, 20130097110001) and a grant from Educational Ministry of China for Basic Scientific Research of National Universities (KYZ201202-9) AbstractIdentifying marker loci related to combining ability (CA) for yield trait in parents of japonica hybrid rice facilitates improving CA of parents and enhancing standard heterosis degree of japonica rice by using molecular marker-assisted selection techniques. F1 seeds of 90 combinations were made by hand-crossed nine CMS lines with ten restorer lines using North Carolina Design II. The F1 populations were planted in Nanjing and Xuyi environments, and six yield traits were investigated. CA of the 19 parental lines was analyzed for six yield traits respectively using the data of 90 F1’s. Combining the data of CA and SSR marker genotypes of the 19 parental lines, SSR marker loci related to CA for six yield traits were detected. Results showed that BT-18A and Wuqiang A were elite CMS lines, and C418 was elite restorer lines in both environments. Number of detected SSR marker loci related to CA for effective panicles per plant, spikelets per panicle, filled grains per panicle, seed setting rate, 1000-grain weight and daily yield per plant were 8, 13, 11, 6, 6, and 2, respectively in Nanjing, 12, 21, 8, 15, 10, and 7, respectively in Xuyi, and 4, 11, 5, 3, 5, and 1, respectively in both environments. Heterozygous genotype marker loci showing positive heterosis accounted for 74% (34/46) in Nanjing, and 53% (39/73) in Xuyi. Among the SSR marker loci detected in both environments, three were each co-associated with CA for three yield traits, and another three for two yield traits. The remaining 14 marker loci were each associated with CA for one yield trait. Through data-base searching, genes/QTLs for the corresponding traits were found nearly ten of the marker loci detected in both environments. Strategies of enhancing CA for yield traits of restorer lines in japonica rice using the marker loci identified in this study were discussed.
Keyword:Hybrid Japonicarice; Backbone parents; Combining ability for yield traits; Marker locus genotypes Show Figures Show Figures
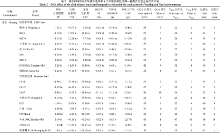
表1 两个环境下90个F1组合6个产量相关性状的表现 Table 1 Performance of six yield-related traits of 90 F1 in two environments
环境 Environment
性状 Trait
最大值 Maximum
最小值 Minimum
平均值 Average
标准差 SD
变异系数 CV (%)
南京 Nanjing
单株有效穗数 PP
12.30
4.70
7.11
1.548
21.8
每穗总粒数 TSP
351.70
145.15
251.99
41.907
16.6
每穗实粒数 FSP
272.50
112.30
193.23
39.339
20.4
结实率 SF (%)
90.90
51.24
76.91
0.091
11.8
千粒重 1000-GW (g)
28.96
21.75
24.97
1.462
5.9
单株日产量 DYP (g)
0.266
0.103
0.160
0.030
18.9
盱眙 Xuyi
单株有效穗数 PP
16.90
6.90
9.82
1.966
20.0
每穗总粒数 TSP
378.80
183.30
282.34
45.827
16.2
每穗实粒数 FSP
303.50
126.50
210.23
35.863
17.1
结实率SF(%)
90.38
45.85
75.35
0.096
12.7
千粒重1000-GW (g)
30.32
23.22
26.37
1.429
5.4
单株日产量 DYP (g)
0.353
0.125
0.237
0.045
19.1
PP: panicles per plant; TSP: total spikelet per panicle; FSP: filled spikelets per panicle; SF: spikelet fertility; 1000-GW: 1000-grain weight; DYP: daily yield per plant. SD: standard deviation. CV: coefficient of variation.
表1 两个环境下90个F1组合6个产量相关性状的表现 Table 1 Performance of six yield-related traits of 90 F1 in two environments
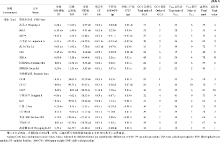
表3 南京环境下检测到的与亲本单个性状配合力显著相关的SSR标记位点 Table 3 Marker loci significantly related to combining ability for each trait in parents in Nanjing environment
标记位点-杂合基因型a Marker locus-heterozygous genotype a
杂合组Heterozygous group
纯合组Homozygous group
位点杂合基因型优势b Heterosis of heterozygous genotype locus b(%)
t值 t-value
v'值c v'-value c
F1组合数Number of F1 cross
各F1组合 平均数 Average of F1’ s
F1组合数 Number of F1 cross
各F1组合 平均数 Average of F1’ s
单株有效穗数 PP
RM16-180/190(3)
36
6.3
54
7.6
-17.4
-4.38
88
RM340-110/160(6)
45
6.4
45
7.8
-17.8
-4.73
73
RM6863-185/195(8)
36
6.3
54
7.6
-17.4
-4.38
88
RM6976-250/280(8)
36
6.3
54
7.6
-17.4
-4.38
88
RM167-150/160(11)
29
8.0
61
6.7
19.1
3.96
43
RM144-230/260(11)
27
8.0
63
6.7
18.6
3.76
43
RM144-240/260(11)
24
6.1
66
7.5
-17.6
-3.81
80
RM247-170/180(12)
24
8.4
66
6.6
26.3
5.44
30
每穗总粒数 TSP
RM3453-155/160(1)
28
275.6
62
241.3
14.2
3.87
71
RM1211-170/180(2)
45
267.6
45
236.4
13.2
3.78
85
RM263-175/180(2)
45
237.1
45
266.9
-11.2
-3.59
87
RM208-180/185(2)
29
225.8
61
264.4
-14.6
-4.51
50
RM5474-170/175(3)
18
284.9
72
243.8
16.9
4.03
29
RM16-180/190(3)
36
279.0
54
234.0
19.2
5.84
79
RM570-200/280(3)
36
271.6
54
238.9
13.7
3.90
81
RM340-110/160(6)
45
274.1
45
229.9
19.2
5.86
87
RM345-180/190(6)
63
266.4
27
218.4
22.0
5.83
43
RM8263-190/195(7)
28
277.9
62
240.3
15.7
4.32
62
RM10-170/180(7)
27
280.2
63
239.9
16.8
4.65
56
RM152-170/175(8)
21
280.1
69
243.4
15.1
3.76
34
RM167-140/150(11)
15
292.3
75
243.9
19.8
4.50
25
每穗实粒数 FSP
RM3453-155/160(1)
28
213.6
62
184.2
16.0
3.49
62
RM259-170/180(1)
37
209.6
53
182.0
15.2
3.48
71
RM341-170/195(1)
35
219.6
55
176.6
24.3
5.94
78
RM208-180/185(2)
29
165.1
61
206.8
-20.2
-5.38
61
RM16-180/190(3)
36
210.5
54
181.9
15.7
3.59
64
RM570-200/280(3)
36
211.5
54
181.2
16.7
3.85
66
RM340-110/160(6)
45
211.1
45
175.6
20.2
4.77
86
RM8263-190/195(7)
28
218.0
62
182.2
19.6
4.38
54
RM6863-185/195(8)
36
210.5
54
181.9
15.7
3.59
64
RM6976-250/280(8)
36
210.5
54
181.9
15.7
3.59
64
RM5652-190/200(9)
19
165.6
71
200.7
-17.5
-3.69
31
结实率 SF
RM341-170/195(1)
35
0.815
55
0.740
10.1
4.12
84
RM406-145/160(2)
63
0.795
27
0.710
11.9
4.46
39
RM574-165/170(5)
30
0.716
60
0.796
-10.1
-4.33
42
RM587-200/230(6)
28
0.823
62
0.745
10.3
4.00
88
RM11-130/150(7)
27
0.720
63
0.790
-8.9
-3.57
38
RM258-150/165(10)
27
0.825
63
0.745
10.6
4.12
87
千粒重 1000-GW
RM406-145/160(2)
63
24.48
27
26.11
-6.3
-5.65
54
RM16-180/190(3)
36
25.68
54
24.49
4.9
4.13
83
RM8263-190/195(7)
28
25.73
62
24.62
4.5
3.54
56
RM10-170/180(7)
27
25.84
63
24.59
5.1
4.00
52
RM6863-185/195(8)
36
25.68
54
24.49
4.9
4.13
83
RM6976-250/280(8)
36
25.68
54
24.49
4.9
4.13
83
单株日产量 DYP
RM208-180/190(2)
21
0.182
69
0.154
17.8
3.21
24
RM406-145/160(2)
63
0.168
27
0.144
16.2
2.94
69
a: RM16-180/190 means that a 180 bp band and a 190 bp band were obtained from female DNA template and male DNA template, respectively, when using RM16 as primer. The digits in parenthesis represent number of chromosome. b: Heterosis of heterozygous locus (%) = (average of trait value in F1heterozygous locus group - average of trait value in F1homozygous group)/ average of trait value in F1homozygous group × 100. c: v': effective degree of freedom. PP: panicles per plant; TSP: total spikelet per panicle; FSP: filled spikelets per panicle; SF: spikelet fertility; 1000-GW: 1000-grain weight; DYP: daily yield per plant. a: RM16-180/190表示用SSR引物RM16扩增母本DNA和父本DNA分别得到180 bp和190 bp条带, 余类推。括号内数字表示染色体号。b: 位点杂合优势(%) = (F1位点杂合组性状平均值 - F1位点纯合组性状平均值)/F1位点纯合组性状平均值× 100。c: v'值表示有效自由度。
表3 南京环境下检测到的与亲本单个性状配合力显著相关的SSR标记位点 Table 3 Marker loci significantly related to combining ability for each trait in parents in Nanjing environment
万建民. 中国水稻遗传育种与品种系谱. 北京: 中国农业出版社, 2010. pp 548-604Wan JM. Genetics and Breeding, Cultivar Pedigree of Rice in China. Beijing: China Agriculture Press, 2010. pp 548-604(in Chinese)[本文引用:2]
[2]
Ma GH, Yuan LP. Hybrid rice achievements and development in China. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 247-256[本文引用:1]
[3]
斯华敏, 刘文真, 付亚萍, 孙宗修, 胡国成. 我国两系杂交水稻发展的现状和建议. 中国水稻科学, 2011, 25: 544-552Si HM, Liu WZ, Fu YP, Sun ZX, Hu GC. Current situation and suggestions for development of two-line hybrid rice in China. Chin J Rice Sci, 2011, 25: 544-552 (in Chinese with English abstract)[本文引用:1]
杜永林, 张巫军, 吴晓燃, 李刚华, 王绍华, 刘正辉, 唐设, 丁艳锋. 江苏省水稻产量时空变化特征. 南京农业大学学报, 2014, 37(5): 7-12. DOI: DOI: 107685/j. issn. 1000-2030. 2014. 05. 002Du YL, Zhang WJ, Wu XR, Li GH, Wang SH, Liu ZH, TangS, Ding YF. The characteristics of spatial and temporal change of rice yield in Jiangsu Province. J Nanjing Agric Univ, 2014, 37(5): 7-12 (in Chinese with English abstract)[本文引用:1]
[6]
邓华凤, 何强, 舒服, 张武汉, 杨飞, 荆彦辉, 东丽, 谢辉. 中国杂交粳稻研究现状与对策. 杂交水稻, 2006, 21(1): 1-6Deng HF, HeQ, ShuF, Zhang WH, YangF, Jing YH, DongL, XieH. Status and technical strategy on development of japonica hybrid rice in China. Hybrid Rice, 2006, 21(1): 1-6 (in Chinese with English abstract)[本文引用:2]
[7]
汤述翥, 张宏根, 梁国华, 严长杰, 刘巧泉, 顾铭洪. 三系杂交粳稻发展缓慢的原因及对策. 杂交水稻, 2008, 23(1): 1-5Tang SZ, Zhang HG, Liang GH, Yan CJ, Liu QQ, Gu MH. Reasons and countermeasures of slow development on three-line japonica hybrid rice. Hybrid Rice, 2008, 23(1): 1-5 (in Chinese with English abstract)[本文引用:1]
[8]
王才林. 江苏省杂交粳稻育种的现状、问题与对策. 西南农业学报, 2009, 24: 1165-1169Wang CL. Current status and prospects of japonica hybrid rice breeding in Jiangsu, China. Southwest China J Agric Sci, 2009, 24: 1165-1169 (in Chinese with English abstract)[本文引用:1]
[9]
杨振玉. 我国杂交粳稻的发展及其技术策略. 杂交水稻, 2005, 20(2): 5-6Yang ZY. Development and technical strategy of japonica hybrid rice in China. Hybrid Rice, 2005, 20(2): 5-6 (in Chinese with English abstract)[本文引用:2]
[10]
洪德林, 汤玉庚. 粳稻雄性不育恢复基因研究: I. 粳稻雄性不育恢复基因的地理分布. 江苏农业学报, 1985, 1(4): 1-5Hong DL, Tang YG. Studies on fertility restoring genes for CMS Oryza sativa SUBSP. KENG in hybrid breeding: I. Geographical distribution of fertility restoring genes for CMS Oryza sativa SUBSP. KENG. J Agric Sci, 1985, 1(4): 1-5 (in Chinese with English abstract)[本文引用:1]
[11]
中国农业科学院, 湖南省农业科学院. 中国杂交水稻的发展. 北京: 农业出版社, 1991. pp 28-40Chinese Academy of Agricultural Sciencesand Hunan Academy of Agricultural Sciences. Development of Hybrid Rice in China. Beijing: Agriculture Press, 1991. pp 28-40(in Chinese)[本文引用:1]
[12]
Gontcharova IK, Gontcharov SV. Hybrid rice breeding in Russia. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 321-327[本文引用:1]
[13]
Bastawisi AO, El-Mowafi H F, Abo Yousef M I, Draz A E, Aidy I R, Maximos M R, Badawi A T. Hybrid research and development in Egypt. In: Virmani S S, Mao C X, Hardy B, eds. Hybrid Rice for Food Security, Poverty Alleviation, and Environmental Protection. Los Baños (Philippines): International Rice Research Institute, 2003. pp 257-263[本文引用:1]
[14]
蔡健, 范海燕, 廖秋平, 马同富, 张桂权. 水稻恢复基因Rf3和Rf4聚合效应分析. 南京农业大学学报, 2014, 37(3): 20-26CaiJ, Fan HY, Liao QP, Ma TF, Zhang GQ. Analysis of pyramiding effect of the Rf3 and Rf4 genes on fertility restoration in rice with cytoplasmic male sterility. J Nanjing Agric Univ, 2014, 37(3): 20-26 (in Chinese with English abstract)[本文引用:1]
[15]
杨振玉, 张宗旭, 魏耀林, 赵迎春, 高勇. 粳型特异亲和恢复系C418的选育及其特性. 杂交水稻, 1998, 13(3): 31-32Yang ZY, Zhang ZX, Wei YL, Zhao YC, GaoY. Breeding and characteristics of japonica type wide compatibility line C418. Hybrid Rice, 1998, 13(3): 31-32 (in Chinese with English abstract)[本文引用:1]
[16]
Liu XC, Wu JL. SSR heterogenic patterns of parents for marking and predicting heterosis in rice breeding. Mol Breed, 1998, 4: 263-268[本文引用:1]
[17]
Liu XC, IshikiK, Wang WX. Identification of favorable AFLP markers to heterosis in hybrid rice. Breed Sci, 2002, 52: 201-206[本文引用:1]
[18]
刘小川, 王渭霞, 陈深广, 余柳青. 杂交稻米质性状的亲本配合力分子标记鉴定. 中国水稻科学, 2005, 19: 25-28Liu XC, Wang WX, Chen GS, Yu LQ. Identification of molecular markers referred to the grain quality characters of rice hybrids based on the combining ability of their parental lines. Chin J Rice Sci, 2005, 19: 25-28 (in Chinese with English abstract)[本文引用:1]
[19]
Liu XC, Chen SC, Chen JS, IshikiK, Wang WX, Yu LQ. Improvement of combining ability for restorer lines with identified SSR marker in hybrid rice breeding. Breed Sci, 2004, 54: 341-346[本文引用:3]
[20]
梁奎, 黄殿成, 赵凯铭, 阮方松, 谢辉, 马文霞, 洪德林. 杂交粳稻亲本产量性状优异配合力的标记基因型筛选. 作物学报, 2010, 36: 1270-1279LiangK, Huang DC, Zhao KM, Ruan FS, XieH, Ma WX, Hong DL. Screening marker genotypes with elite combining ability for yield traits in parents of hybrid japonica rice (Oryza sativa L. ). Acta Agron Sin, 2010, 36: 1270-1279 (in Chinese with English abstract)[本文引用:4]
[21]
黄殿成, 梁奎, 孙程, 江建华, 石明亮, 戴剑, 谢辉, 赵凯铭, 阮方松, 马文霞, 洪德林. 杂交粳稻亲本米质性状优异配合力的标记基因型鉴定. 作物学报, 2011, 37: 405-414Huang DC, LiangK, SunC, Jiang JH, Shi ML, DaiJ, XieH, Zhao KM, Ruan FS, Ma WX, Hong DL. Identification of marker genotypes associated with elite combining ability for quality traits in parents of hybrid japonica rice (Oryza sativa L. ). Acta Agron Sin, 2011, 37: 405-414 (in Chinese with English abstract)[本文引用:2]
[22]
黄殿成, 阮方松, 刘健, 谢辉, 赵凯铭, 梁奎, 江建华, 蔡勇林, 洪德林. 杂交粳稻亲本穗长和枝梗数性状优异配合力的标记基因型筛选. 南京农业大学学报, 2011, 34(2): 1-7Huang DC, Nguyen PT, LiuJ, XieH, Zhao KM, LiangK, Jiang JH, Cai YL, Hong DL. Screening marker genotypes with elite combining ability for panicle length and branch numbers in parents of hybrid japonica rice (Oryza sativa L. ). J Nanjing Agric Univ, 2011, 34(2): 1-7 (in Chinese with English abstract)[本文引用:2]
[23]
刘二宝, 刘洋, 刘晓丽, 刘强明, 赵凯铭, Edzesi WM, 洪德林. 杂交粳稻亲本产量性状优异配合力的标记基因型检测. 中国水稻科学, 2013, 27: 473-481Liu EB, LiuY, Liu XL, Liu QM, Zhao KM, Edzesi WM, Hong DL. Detecting marker genotypes with elite combining ability for yield traits in parents of hybrid japonica rice. Chin J Rice Sci, 2013, 27: 473-481 (in Chinese with English abstract)[本文引用:4]
[24]
刘洋, 刘二宝, 曾思远, 刘强明, 普伟, 梁利君, 刘佩睿, 张琪梦, 洪德林. 杂交粳稻亲本米质性状配合力的分子标记区段检测. 中国水稻科学, 2015, 29: 443-451LiuY, Liu EB, Zeng SY, Liu QM, PuW, Liang LJ, Liu PR, Zhang QM, Hong DL. Identification of molecular marker fragments associated with combining ability for quality traits in parents of hybrid japonica rice (Oryza sativa L. ). Chin J Rice Sci, 2015, 29: 443-451 (in Chinese with English abstract)[本文引用:2]
[25]
程宝山, 万志兵, 洪德林. 35个粳稻品种SSR指纹图谱的构建及遗传相似性分析. 南京农业大学学报, 2007, 30(3): 1-8Cheng BS, Wan ZB, Hong DL. Establishment of SSR finger print map and analysis of genetic similarity among 35 varieties in japonica rice (Oryza sativa L. ). J Nanjing Agric Univ, 2007, 30(3): 1-8 (in Chinese with English abstract)[本文引用:1]
[26]
莫惠栋. p×q交配模式的配合力分析. 江苏农学院学报, 1982, 3(3): 51-57Mo HD. The analysis of combining ability in p×q mating pattern (continued). J Jiangsu Agric Coll, 1982, 3(3): 51-57 (in Chinese)[本文引用:1]
[27]
莫惠栋. p×q交配模式的配合力分析(续). 江苏农学院学报, 1982, 3(4): 53-57Mo HD. The analysis of combining ability in p×q mating pattern (continued). J Jiangsu Agric Coll, 1982, 3(4): 53-57 (in Chinese)[本文引用:1]
[28]
徐辰武. 稻米品质性状的遗传研究. 南京农业大学博士学位论文, 江苏南京, 1998Xu CW. Inheritance of Quality Characters of Rice Grain. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 1998 (in Chinese with English abstract)[本文引用:2]
[29]
Russell WA. Genetic improvement of maize yield. Adv Agron, 1991, 46: 245-298[本文引用:1]
[30]
Dudly JW, Saghai MA, Rufener GK. Molecular marker and grouping of parents in maize breeding programs. Crop Sci, 1991, 31: 718-723[本文引用:1]
[31]
王胜军, 万建民, 陆作楣. 利用SSR分子标记划分杂交籼稻亲本群的研究. 作物学报, 2006, 32: 1437-1443Wang SJ, Wan JM, Lu ZM. Parental cluster analysis in indica hybrid rice (Oryza sativa L. ) by SSR analysis. Acta Agron Sin, 2006, 32: 1437-1443 (in English with Chinese abstract)[本文引用:1]
[32]
王胜军, 陆作楣. 我国常用杂交籼稻亲本杂种优势群的初步研究. 南京农业大学学报, 2007, 30(1): 14-18Wang SJ, Lu ZM. Study on heterosis groups of key parental lines in indica hybrid rice (Oryza sativa L. ) in China. J Nanjing Agric Univ, 2007, 30(1): 14-18 (in Chinese with English abstract)[本文引用:1]
[33]
Virmani SS, Aquino RC, Khush GS. Heterosis breeding in rice (Oryza sativa L. ). Theor Appl Genet, 1982, 63: 373-380[本文引用:1]
[34]
Huang XH, Yang SH, Gong JY, ZhaoY, FengQ, GongH, Li WJ, Zhan QL, Cheng BY, Xia JH, ChenN, Hao ZN, Liu KY, Zhu CR, HuangT, ZhaoQ, ZhangL, Fan DL, Zhou CC, Lu YQ, Weng QJ, Wang ZX, Li JY, Han, B. Genomic analysis of hybrid rice varieties reveals numerous superior alleles that contribute to heterosis. Nat Commun, 2015, 6: 6258, DOI: DOI:10.1038/ncomms7258[本文引用:1]



{kind=link}
{kind=link}