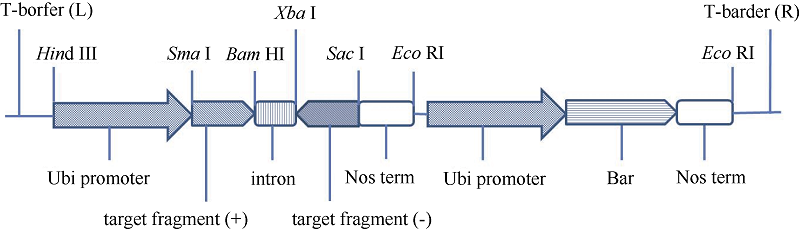
关键词:小麦1B/1R易位系; 黑麦碱; RNA干扰; 基因沉默; 加工品质 Effect of ω-Secalin Gene Silencing on Processing Quality of Wheat 1B/1R Translocation Line CHAI Jian-Fang*, WANG Hai-Bo*, MA Xiu-Ying, ZHANG Cui-Mian, DONG Fu-Shuang Genetics and Physiology Institute, Hebei Academy of Agriculture and Forestry Sciences / Plant Genetic Engineering Center of Hebei Province, Shijiazhuang 050051, China Fund:This study was supported by the Natural Science Foundation of Hebei Province, China (C2014301005) AbstractOmega-secalin is an important factor for the poor processing quality of wheat 1B/1R translocation lines. In this study, an expression vector of silencing ω-secalin gene was constructed and transferred into wheat variety Jinhe 9123 via Agrobacterium tumefaciens strain EH105. The three T0 transgenic plants obtained were consecutively multiplied after PCR validation until the T4 lines. Acid-PAGE assay indicated that total amount of ω-secalins in the transgenic lines decreased 53% on average compared with that in their respective control. In the three homozygous transgenic lines, gluten index, precipitation value, and stabilization time increased significantly, whereas the agronomic traits, such as plant height, grain number per spike, 1000-grain weight and plot yield, had no significant changes compared with those in the wild type. These results suggest that silencing ω-secalin genes can improve the processing quality of wheat 1B/1R translocation lines on the premise of not affecting grain yield.
Keyword:Wheat 1B/1R translocation line; Secalin; RNAi; Gene silencing; Processing quality Show Figures Show Figures
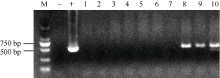
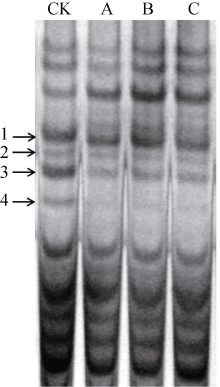
表1 转基因小麦株系及其受体对照种子中不同ω -黑麦碱的表达量比较 Table 1 Comparison of ω -secalins in seeds of wheat transgenic lines and the non-transgenic recipient
ω 黑麦碱条带 ω -secalin band
受体对照 Recipient control (%)
转基因株系 Transgenic lines (%)
转基因株系/受体对照比值 Transgenic lines/recipient control ratio
1
43.2
24.6*
0.569
2
4.8
4.8
0.995
3
36.4
15.4* *
0.424
4
15.6
2.7* *
0.170
合计Total
100.0
47.5
* and * * indicate significant difference between the transgenic line and the recipient control at P < 0.05 and P < 0.01, respectively. * 和* * 分别表示转基因株系与受体对照之间在P < 0.05和P < 0.01水平差异显著。
表1 转基因小麦株系及其受体对照种子中不同ω -黑麦碱的表达量比较 Table 1 Comparison of ω -secalins in seeds of wheat transgenic lines and the non-transgenic recipient
周阳, 何中虎, 张改生, 夏兰琴, 陈新民, 高永超, 井赵斌, 于广军. 1BL/1RS易位系在我国小麦育种中的应用. 作物学报, 2004, 30: 531-535ZhouY, He ZH, Zhang GS, Xia LQ, Chen XM, Gao YC, Jing ZB, Yu GJ. Utilization of 1BL/1RS translocation in wheat breeding in China. Acta Agron Sin, 2004, 30: 531-535 (in Chinese with English abstract)[本文引用:1]
[2]
杨足君, 任正隆. 抗白粉病基因Pm8在四川小麦中遗传表达初步研究. 四川农业大学学报, 1997, 15: 452-456Yang ZJ, Ren ZL. Expression of gene Pm8 for resistance to powdery mildew in wheat for Sichuan. J Sichuan Agric Univ, 1997, 15: 452-456 (in Chinese with English abstract)[本文引用:1]
[3]
Shi ZX, Chen XM, Line RF, LeungH, Wellings CR. Development of resistance gene analog polymorphism markers for the Yr9 gene resistance to wheat stripe rust. Genome, 2001, 44: 509-516[本文引用:1]
[4]
Lei MP, Li GR, LiuC, Yang ZJ. Characterization of wheat-Secale africanum introgression lines reveals evolutionary aspects of chromosome 1R in rye. Genome, 2012, 55: 765-774[本文引用:1]
[5]
Yang MY, Ren TH, Yan BJ, LiZ, Ren ZL. Diversity resistance to Puccinia striiformis f. sp tritici in rye chromosome arm 1RS expressed in wheat. Genet Mol Res, 2014, 13: 8783-8793[本文引用:1]
[6]
Dhaliwal AS, Mares DJ, Marshall DR. Measurement of dough surface stickness associated with 1B/1R chromosome translocation in bread wheats. Cereal Sci, 1990, 12: 165-175[本文引用:1]
[7]
刘建军, 何中虎, Peña RJ, 赵振东. 1BL/1RS易位对小麦加工品质的影响. 作物学报, 2004, 30: 149-153Liu JJ, He ZH, Peña RJ, Zhao ZD. The effects of 1B/1R translocation on grain quality and noodle quality of bread wheat. Acta Agron Sin, 2004, 30: 149-153 (in Chinese with English abstract)[本文引用:1]
[8]
Graybosh RA, Peterson CJ, Hansen LE, Mattern PJ. Relationships between protein solubility characteristics, 1BL/1RS, high molecular weight glutenin composition, and end-use quality in winter wheat germ plasm. Cereal Chem, 1990, 67: 342-349[本文引用:1]
[9]
Lukaszewski AJ. Manipulation of the 1RS·1BL translocation in wheat by induced homoeologous recombination. Crop Sci, 2000, 40: 216-225[本文引用:3]
[10]
FuS, TangZ, RenZ, ZhangH. Transfer to wheat (Triticum aestivum) of small chromosome segments from rye (Secale cereale) carrying disease resistance genes. J Appl Genet, 2010, 51: 115-121[本文引用:3]
[11]
Scott MH, Amy AC, Gregory JH. Post-transcriptional gene silencing by double-strand ed RNA. Nat Rev Genet, 2001, 2: 110-119[本文引用:1]
Li JR, ZhaoW, Li QZ, Ye XG, An BY, LiX, Zhang XS. RNA silencing of waxy gene results in low levels of amylose in the seeds of transgenic wheat (Triticum aestivum L. ). Acta Genet Sin, 2005, 32: 846-854[本文引用:1]
[14]
孙重霞, 杨凤萍, 张婷, 隋晓燕, 梁荣奇, LiuQ, 张晓东, 李保云. 利用RNAi技术抑制籽粒PPO合成改良小麦面粉白度的研究. 中国农业科学, 2013, 46: 1104-1113Sun CX, Yang FP, ZhangT, Sui XY, Liang RQ, LiuQ, Zhang XD, Li BY. Down-regulation of the expression of grain ppo genes to improve wheat dough whiteness by RNA interference. Sci Agric Sin, 2013, 46: 1104-1113 (in Chinese with English abstract)[本文引用:1]
[15]
Chai JF, LiuX, Jia JZ. Homoeologous cloning of ω-secalin gene family in a wheat 1BL/1RS translocation. Cell Res, 2005, 15: 658-664[本文引用:3]
[16]
MuraiJ, TairaT, OhtaD. Isolation and characterization of the three Waxy genes encoding the granule-bound starch synthase in hexaploid wheat. Gene, 1999, 234: 71-79[本文引用:1]
[17]
张学勇, 杨欣明, 董玉琛. 醇溶蛋白电泳在小麦种质资源遗传分析中的应用. 中国农业科学, 1995, 28(4): 25-32Zhang XY, Yang XM, Dong YC. Genetic analysis of wheat germplasm by acid polyacrylamide gel electrophoresis of gliadins. Sci Agric Sin, 1995, 28(4): 25-32 (in Chinese with English abstract)[本文引用:1]
[18]
纪军, 刘冬成, 王静, 李俊明, 张爱民. 一种小麦高、低分子量麦谷蛋白亚基的提取方法. 遗传, 2008, 30: 123-126JiJ, Liu DC, WangJ, Li JM, Zhang AM. A method of extraction and separation of wheat gluten. Hereditas (Beijing), 2008, 30: 123-126 (in Chinese with English abstract)[本文引用:1]
[19]
Gil-HumanesJ, PistónF, TollefsenS, Sollid LM, BarroF. Effective shutdown in the expression of celiac disease-related wheat gliadin T-cell epitopes by RNA interference. Proc Natl Acad Sci USA, 2010, 107: 17023-17028[本文引用:1]
[20]
HussainA, Lukow OM. Characterization of the 1B/1R translocation in wheat using water extractable protein concentrate. Euphytica, 1994, 78: 109-113[本文引用:1]
[21]
SharmaS, Bhat PR, EhdaieB, Close TJ, Lukaszewski AJ, Waines JG. Integrated genetic map and genetic analysis of a region associated with root traits on the short arm of rye chromosome 1 in bread wheat. Theor Appl Genet, 2009, 119: 783-793[本文引用:1]
[22]
相微微, 张怀刚, 王道文, 柳觐, 刘宝龙. 转基因小麦沉默的HMW-GS基因的遗传及表达. 麦类作物学报, 2009, 29: 185-188Xiang WW, Zhang HG, Wang DW, LiuJ, Liu BL. Inheritance and expression of silenced HMW-GS genes in wheat. J Triticeae Crops, 2009, 29: 185-188 (in Chinese with English abstract)[本文引用:1]
[23]
武茹, 高德荣, 别同德, 张晓, 赵芸, 程顺和. 转基因小麦1Dx5基因沉默的遗传表达和品质效应分析. 麦类作物学报, 2010, 30: 991-996WuR, Gao DR, Bie TD, ZhangX, ZhaoY, Cheng SH. Genetic analysis and quality effects of 1Dx5 subunit silent in transgenic wheat. J Triticeae Crops, 2010, 30: 991-996 (in Chinese with English abstract)[本文引用:1]
 , 王海波
, 王海波





{kind=link}
{kind=link}
{kind=link}
{kind=link}