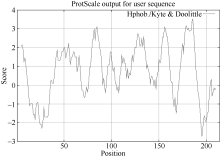
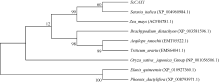
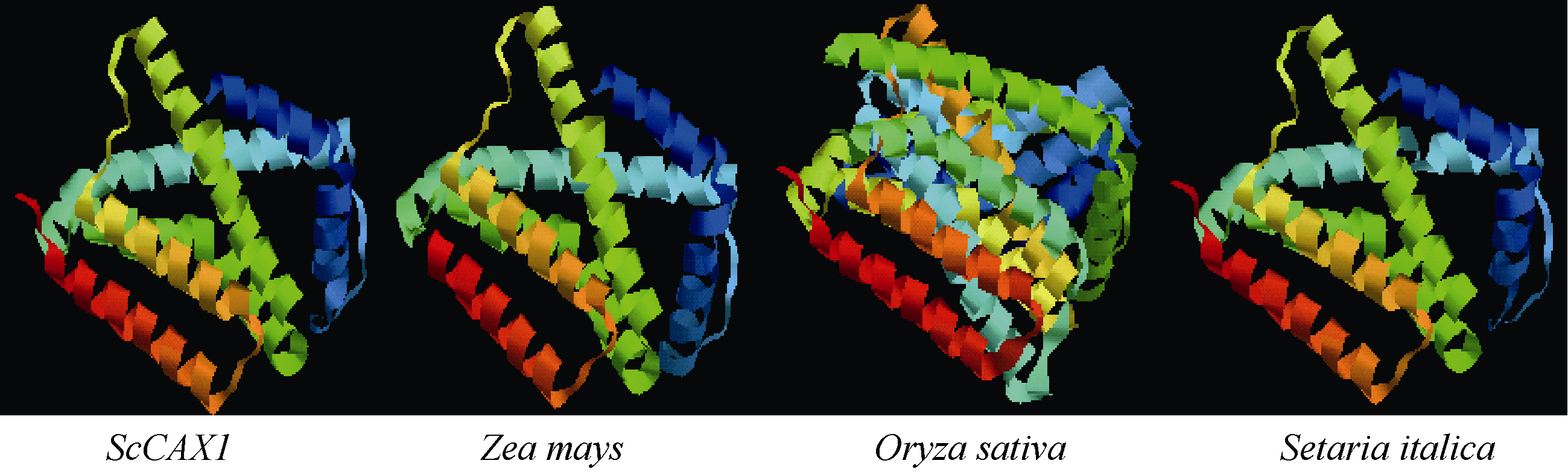
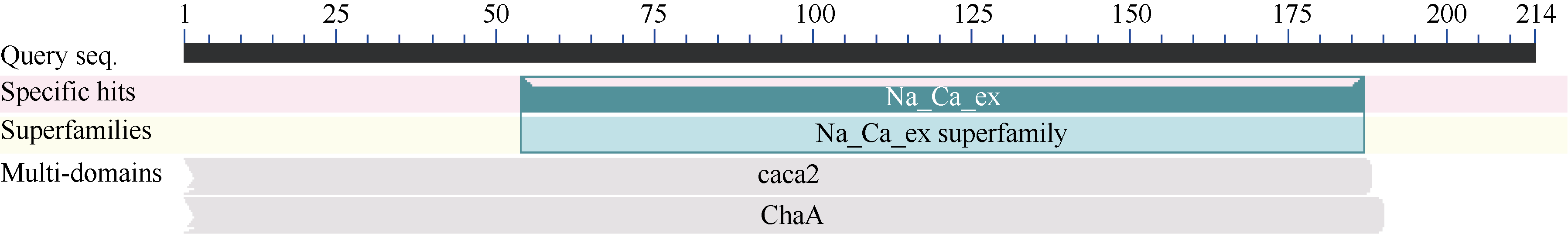
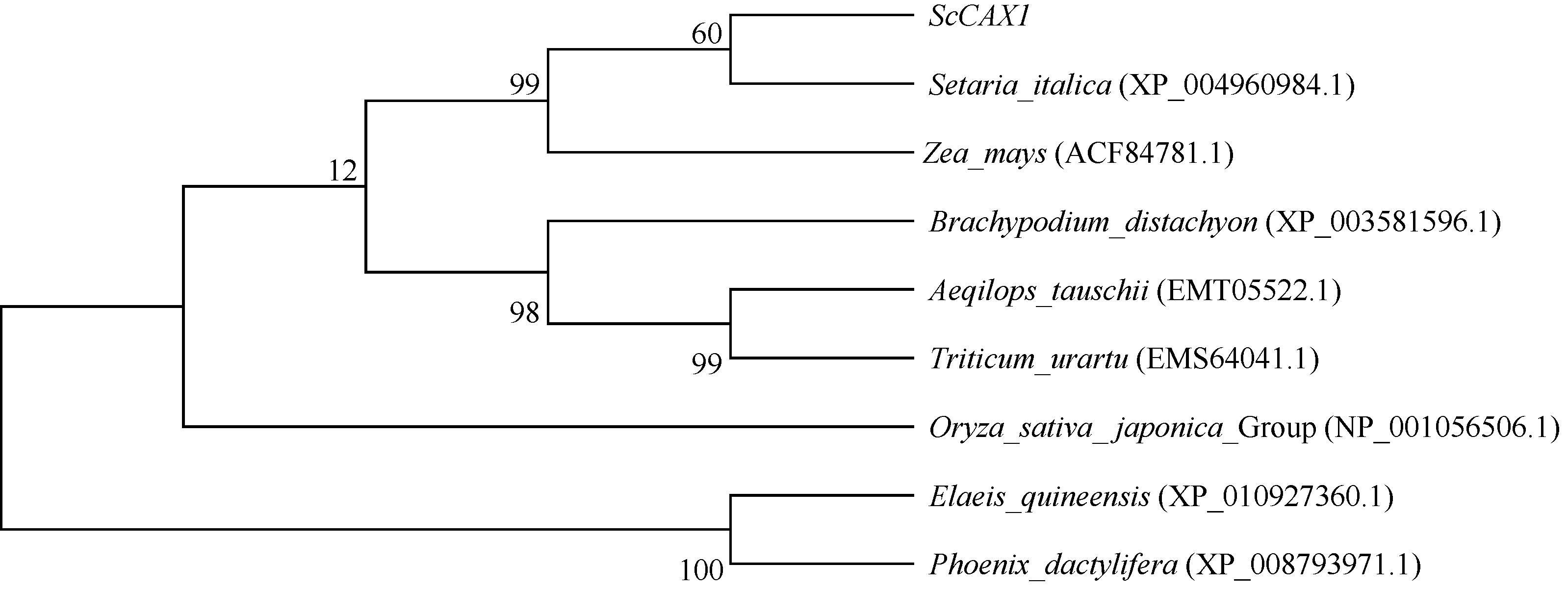
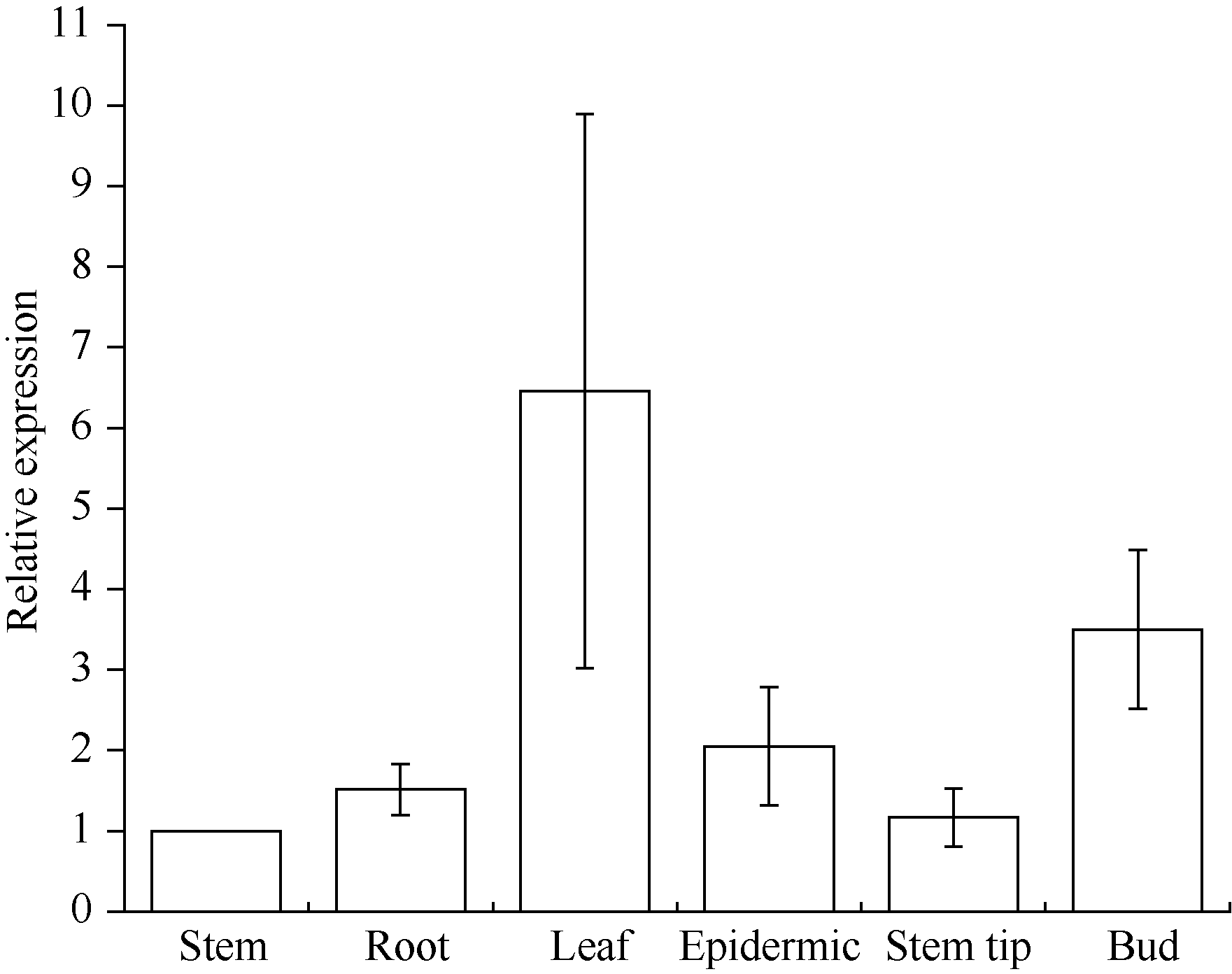
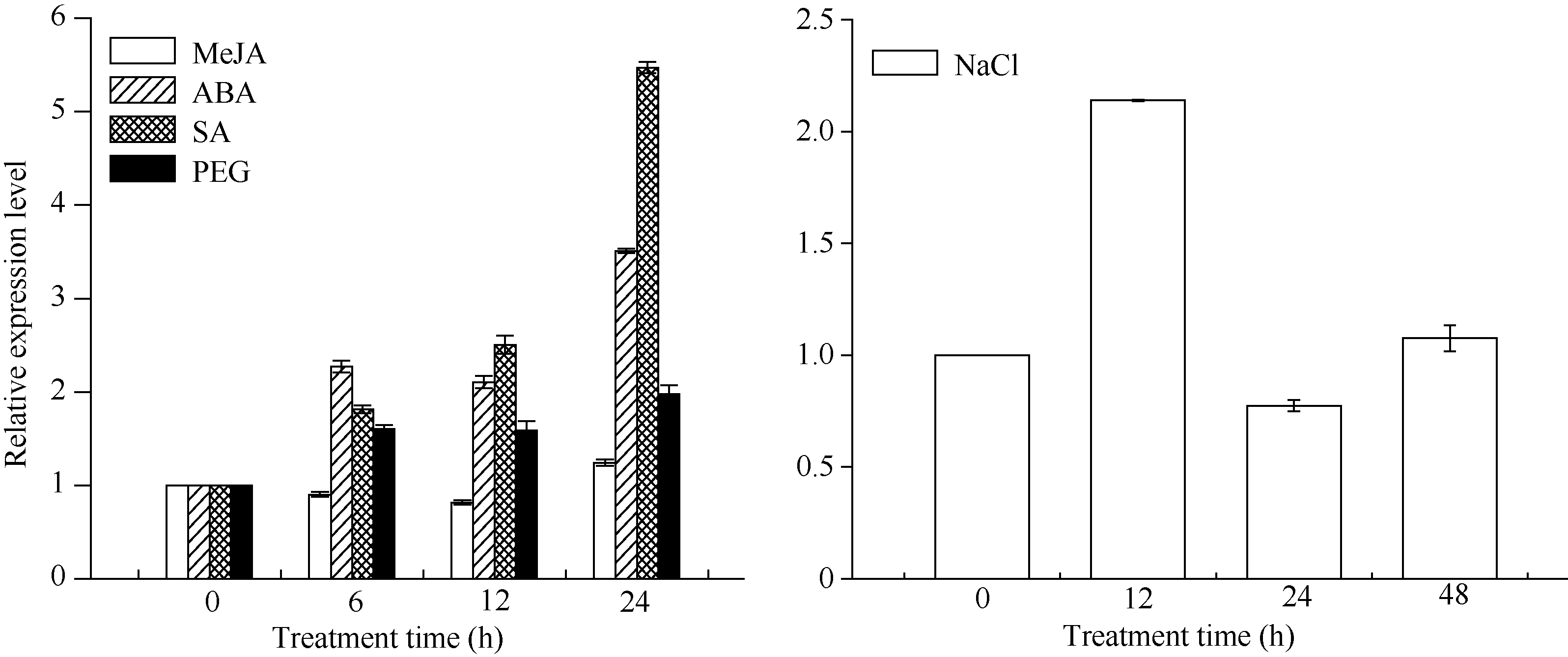
关键词:甘蔗; CAX1基因; 电子克隆; 生物信息学; 实时荧光定量PCR Cloning and Expression Analysis of a Ca2+/H+ Antiporter Gene from Sugarcane SU Wei-Hua**, LIU Feng**, HUANG Long, SU Ya-Chun, HUANG Ning, LING Hui, WU Qi-Bin, ZHANG Hua, QUE You-Xiong* Key Laboratory of Sugarcane Biology and Genetic Breeding (Fujian), Ministry of Agriculture, Fujian Agriculture and Forestry University / Sugarcane Research & Development Center, China Agricultural Technology System, Fuzhou 350002, China Fund:This study was supported by the China Agriculture Research System (CARS-20), the Special Fund for Agro-Scientific Research in the Public Interest (201503119), and the Program for New Century Excellent Talents in Fujian Province University (JA14095) AbstractCAX (Ca2+/H+ antiporter) is a major category of Ca2+ active transport systems in plant cell membrane. In the present study, using a CAX1 mRNA sequence from Sorghum bicolor (GenBank accession number: XM_002441593) as the probe, the full-length cDNA sequence of sugarcane CAX1 gene was cloned by in silico cloning combined with RT-PCR amplification, and named as ScCAX1(GenBank accession number: KT799799). Bioinformatics analysis showed that ScCAX1 has a length of 784 bp and contains a complete open reading frame with a length of 645 bp, which encodes a 214 amino acid residues of sugarcane CAX1 protein. The ScCAX1 protein with stable acidity and hydrophobia was detected to be located in thylakoid membrane of chloroplasts with no signal peptide. It belongs to a conserved Na_Ca_ex. The mainly secondary structure element of ScCAX1 protein is alpha helix. Real time quantitative PCR (RT-qPCR) analysis revealed that the expression of ScCAX1 was tissue-specific, with constituent expression in different tissues of sugarcane. The highest expression was observed in leaf while the lowest in stem. Besides, the expression of ScCAX1gene could be regulated by treatments of PEG, NaCl, SA, ABA, and MeJA. The expression level of this gene was up-regulated by ABA, SA and PEG, with the highest inducible expression level in treatment of 24 hours. The expression level was 5.47 times higher than that of control under 24 hours stress of SA, and 3.5 times higher than that of control under 24 hours stress of ABA. Under 6 hours stress of NaCl, the gene had the highest inducible expression level, which was 2.14 times higher than that of control. This study suggested that ScCAX1 could response to stresses, and its expression may be associated with salt resistance and osmotic tolerance in sugarcane.
Keyword:Sugarcane; CAX1 gene; in silico cloning; Bioinformatics; Real-time quantitative PCR Show Figures Show Figures
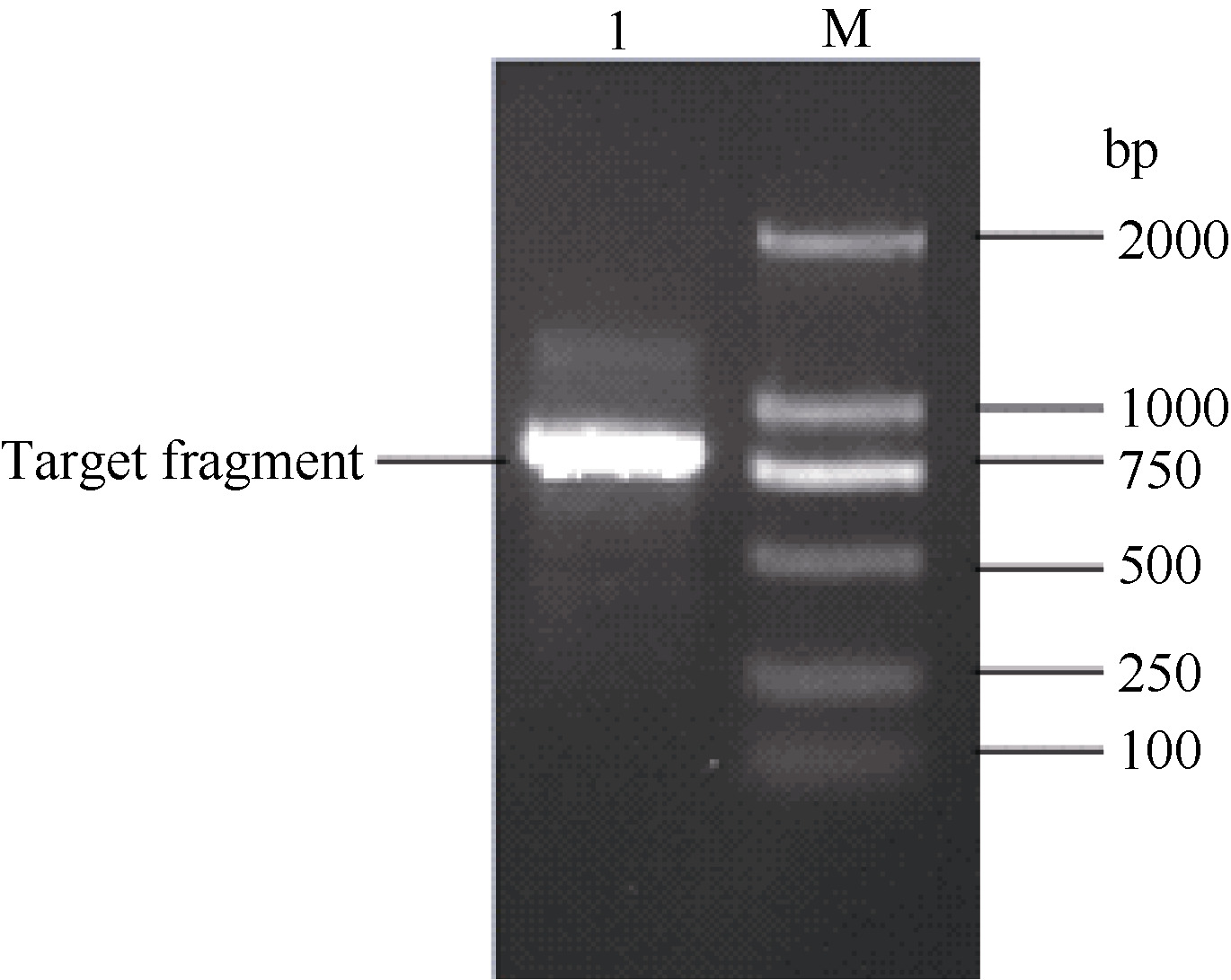
图2 同源克隆获得的甘蔗ScCAX1基因的cDNA序列及其推导的氨基酸序列(* 终止密码子) 黑色框部分为特异性引物在基因序列中的位置。Fig. 2 Nucleotide sequence and deduced amino acid sequence of sugarcane ScCAX1 gene obtained by homology cloning (* stop codon) The sequence fragment complementary to primer is highlighted in black box.
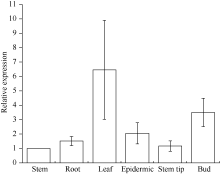
图8 甘蔗ScCAX1基因在不同组织中的表达误差线为每组处理的标准误差(n = 3)。Fig. 8 Relative expression of ScCAX1 gene in different tissues of sugarcaneError bars represent the standard error of each treating group (n = 3).
图9 甘蔗 ScCAX1基因在不同外源胁迫下的表达 误差线为每组处理的标准误差(n = 3)。Fig. 9ScCAX1gene expression in sugarcane under different exogenous stresses Error bars represent the standard error of each treating group (n = 3).
Sand ersD, PellouxJ, BrownleeC, Harper JF. Calcium at the crossroads of signaling. Plant Cell, 2002, 14(suppl): S401-S417[本文引用:2]
[3]
Curran AC, HwangI, CorbinJ, MartinezS, RayleD, SzeH, Harper JF. Autoinhibition of a calmodulin-dependent calcium pump involves a structure in the stalk that connects the transmembrane domain to the ATPase catalytic domain. J Biol Chem, 2000, 275: 30301-30308[本文引用:2]
[4]
祁碧菽, 李春光, 陈叶苗, 陆平利, 郝福顺, 沈国明, 陈珈, 王学臣. 水稻Ca2+/H+反向转运体OsCAX3的功能分析和亚细胞定位研究. 生物化学与生物物理进展, 2005, 32: 876-882Qi BS, Li CG, Chen YM, Lu PL, Hao FS, Shen GM, ChenJ, Wang XC. Functional analysis of rice Ca2+/H+ antiporter OsCAX3 in yeast and its subcellular localization in plant. Prog Biochem Biophys, 2005, 32: 876-882 (in Chinese with English abstract)[本文引用:3]
[5]
朱晓军, 杨劲松, 梁永超, 娄运生, 杨晓英. 盐胁迫下钙对水稻幼苗光合作用及相关生理特性的影响. 中国农业科学, 2004, 37: 1497-1503Zhu XJ, Yang JS, Liang YC, Lou YS, Yang XY. Effects of exogenous calcium on photosynthesis and its related physiological characteristics of rice seedlings under salt stress. Sci Agric Sin, 2004, 37: 1497-1503 (in Chinese with English abstract)[本文引用:1]
[6]
McCormack E, Tsai Y C, Braam J. Hand ling calcium signaling: Arabidopsis CaMs and CMLs. Plant Sci, 2005, 10: 383-389[本文引用:1]
[7]
MahajanS, TutejaN. Calcium signaling network in plants: an overview. Plant Signal Behav, 2007, 2: 79-85[本文引用:2]
[8]
ChinnusamyV, Zhu JK. Plant salt tolerance. Top Curr Genet, 4: 241-270[本文引用:1]
[9]
White PJ, Broadley MR. Calcium in plants. Ann Bot, 2003, 92: 487-511[本文引用:1]
[10]
陈志远. 拟南芥AtCCX1基因的克隆、表达和功能鉴定. 西北农林科技大学博士学位论文, 陕西西安, 2011Chen ZY. Clone, Expression and Characterization of Arabidopsis AtCCX1. PhD Dissertation of North West Agriculture and Forestry University, Xi’an, China, 2011 (in Chinese with English abstract)[本文引用:1]
[11]
Hirschi KD, Zhen RG, Cunningham KW, Rea PA, Fink GR. CAX1, an H+/Ca2+ antiporter from Arabidopsis. Proc Natl Acad Sci USA, 1996, 93: 8782-8786[本文引用:2]
[12]
Hirschi KD, Korenkov VD, Wilganowski NL, Wagner GJ. Expression of Arabidopsis CAX2 in tobacco. Altered metal accumulation and increased manganese tolerance. Plant Physiol, 2000, 124: 125-133[本文引用:1]
[13]
KamiyaT, Akahori TM. Expression profile of the genes for rice cation/H+ exchanger family and functional analysis in yeast. Plant Cell Physiol, 2005, 46: 1735-1740[本文引用:1]
[14]
刘赵越, 童伟, 张英华, 方荣俊, 赵卫国, 李龙. 桑树Ca2+/H+反向转运体基因MCAX1的克隆及序列与表达分析. 蚕业科学, 2012, 38: 192-198Liu ZY, TongW, Zhang YH, Fang RJ, Zhao WG, LiL. Molecular cloning, sequence and expression analyses of Ca2+/H+ antiporter gene in mulberry (Morus L. ). Acta Sericol Sin, 2012, 38: 192-198 (in Chinese with English abstract)[本文引用:1]
[15]
Pittman JK, Hirschi KD. Regulation of CAX1, an Arabidopsis Ca2+/H+ antiporter. Identification of an N-terminal autoinhibitory domain. Plant Physiol, 2001, 127: 1020-1029[本文引用:1]
[16]
Cheng NH, Pittman JK, ShigakiT, Hirschi KD. Characterization of CAX4, an Arabidopsis H+/cation antiporter. Plant Physiol, 2002, 128: 1245-1254[本文引用:1]
[17]
MaserP, ThomineS, Schroeder JI, Ward JM, HirschiK, SzeH, Talke IN, AmtmannA, Maathuis F J M, Sand ers D, Harper J F, Tchieu J, Gribskov M, Persans M W, Salt D E, Kim S A, Guerinot M L. Phylogenetic relationships within cation transporter families of Arabidopsis. Plant Physiol, 2001, 126: 1646-1667[本文引用:1]
[18]
KamiyaT, AkahoriT, AshikariM, MaeshimaM. Expression of the vacuolar Ca2+/H+ exchanger, OsCAX1a, in rice: cell and age specificity of expression, and enhancement by Ca2+. Plant Cell Physiol, 2006, 47: 96-106[本文引用:1]
[19]
Ueoka-NakanishiH, NakanishiY, TanakaY, MaeshimaM. Properties and molecular cloning of Ca2+/H+ antiporter in the vacuolar membrane of mung bean. Eur J Biochem, 1999, 262: 417-425[本文引用:1]
[20]
许莲. 棉花Ca2+转运相关基因的克隆与功能鉴定. 华中农业大学硕士学位论文, 湖北武汉, 2011XuL. Isolation and Characterization of Ca2+ Transport Related Genes in Cotton. MS Thesis of Huazhong Agricultural University, Wuhan, China, 2011 (in Chinese with English abstract)[本文引用:2]
[21]
Cheng NH, Hirschi KD. Cloning and characterization of CXIP1, a novel PICOT domain-containing Arabidopsis protein that associates with CAX1. J Biol Chem, 2003, 278: 6503-6509[本文引用:1]
[22]
Hirschi KD. Expression of Arabidopsis CAX1 in tobacco: altered calcium homeostasis and increased stress sensitivity. Plant Cell, 1999, 11: 2113-2122[本文引用:2]
[23]
ParkS, Cheng NH, Pittman JK, Yoo KS, ParkJ, Smith RH, Hirschi KD. Increased calcium levels and prolonged shelf life in tomatoes expressing Arabidopsis H+/Ca2+ transporters. Plant Physiol, 2005, 139: 1194-1206[本文引用:3]
[24]
ParkS, Kim CK, Pike LM, Smith RH, Hirschi KD. Increased calcium in carrots by expression of an Arabidopsis H+/Ca2+ transporters. Mol Breed, 2004, 14: 275-282[本文引用:1]
[25]
Luo GZ, Wang HW, HuangJ, Tiara AG, Wang YJ, Zhang JS, Chen SY. A putative plasma membrane cation/proton antiporter from soybean confers salt tolerance in Arabidopsis. Plant Mol Biol, 2005, 59: 809-820[本文引用:2]
[26]
Pittman JK, EdmondC, Sunderland PA, Bray CM. A cation-regulated and proton gradient-dependent cation transporter from Chlamydomonas reinhardtii has a role in calcium and sodium homeostasis. J Biol Chem, 2009, 284: 525-533[本文引用:1]
[27]
CatalaR, SantosE, Alonso JM, Ecker JR, Martinez-Zapater J M, Salinas J. Mutations in the Ca2+/H+ transporter CAX1 increase CBF/DREB1 expression and the cold-acclimation response in Arabidopsis. Plant Cell, 2003, 15: 2940-2951[本文引用:1]
[28]
马改艳, 徐学荣. 对当前我国甘蔗产业发展形势的分析与思考. 云南农业大学学报, 2013, 7(6): 29-35Ma GY, Xu XR. Analysis and deliberation on the current development situation of China’s sugar industry. J Yunnan Agric Univ, 2013, 7(6): 29-35 (in Chinese with English abstract)[本文引用:1]
[29]
黄珑, 苏炜华, 张玉叶, 黄宁, 凌辉, 肖新换, 阙友雄, 陈如凯. 甘蔗CIPK基因的同源克隆与表达. 作物学报, 2015, 41: 499-506HuangL, Su WH, Zhang YY, HuangN, LingH, Xiao XH, Que YX, Chen RK. Cloning and expression analysis of CIPK gene in sugarcane. Acta Agron Sin, 2015, 41: 499-506 (in Chinese with English abstract)[本文引用:2]
[30]
GeourjonC, DeleageG. SOPMA: significant improvements in protein secondary structure prediction by consensus prediction from multiple alignments. Comp Appl Biosci, 1995, 11: 681-684[本文引用:1]
[31]
Guo JL, LingH, Wu QB, Xu LP, Xue YX. The choice of reference genes for assessing gene expression in sugarcane under salinity and drought stresses. Sci Rep, 2014, 4: 7042-7042[本文引用:1]
[32]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using Real-time quantitative PCR and the 2-∆∆CT method. Methods, 2001, 25: 402-408[本文引用:1]
[33]
周绚. 苹果Ca2+/H+反向转运体活性及其基因表达特性研究. 南京农业大学硕士学位论文, 江苏南京, 2009ZhouX. Study on Ca2+/H+ antiporter activity and its gene expression of apple. MS Thesis of Nanjing Agricultural University, Nanjing, China, 2009 (in Chinese with English abstract)[本文引用:2]
[34]
Reddy A S N. Calcium: silver bullet in signaling. Plant Sci, 2001, 160: 381-404[本文引用:1]
[35]
MiedemaH, Bothwell JH, BrownleeC, Davies JM. Calcium uptake by plant cells-channels and pumps acting in concert. Trends Plant Sci, 2001, 6: 514-519[本文引用:1]
[36]
刘新, 孟繁霞, 张蜀秋, 娄成后. Ca2+参与水杨酸诱导蚕豆气孔运动时的信号转导. 植物生理与分子生物学学报, 2003, 29: 59-64LiuX, Meng FX, Zhang SQ, Lou CH. Ca2+ is involved in the signal transduction during stomatal movement in Vicia faba L. induced by salicylic acid. J Plant Physiol Mol Biol, 2003, 29: 59-64 (in Chinese with English abstract)[本文引用:1]
[37]
郭园园, 陈江华, 周慧娜, 商慧文, 翟妞, 张艳玲. 不同镉积累基因型烟草CAX2基因克隆及序列分析. 南方农业学报, 2015, 46: 181-187Guo YY, Chen JH, Zhou HN, Shang HW, ZhaiN, Zhang YL. Cloning and sequence analysis of CAX2 from two Nicotiana genotypes with different Cd accumulating pattern. J Southern Agric, 2015, 46: 181-187 (in Chinese with English abstract)[本文引用:1]
[38]
NielsenH, EngelbrechtJ, BrunakS, Heijine GV. Identification of prokaryotic and eukaryotic signal peptides and prediction of their cleavage site. Protein Eng, 1997, 10: 1-6[本文引用:1]
[39]
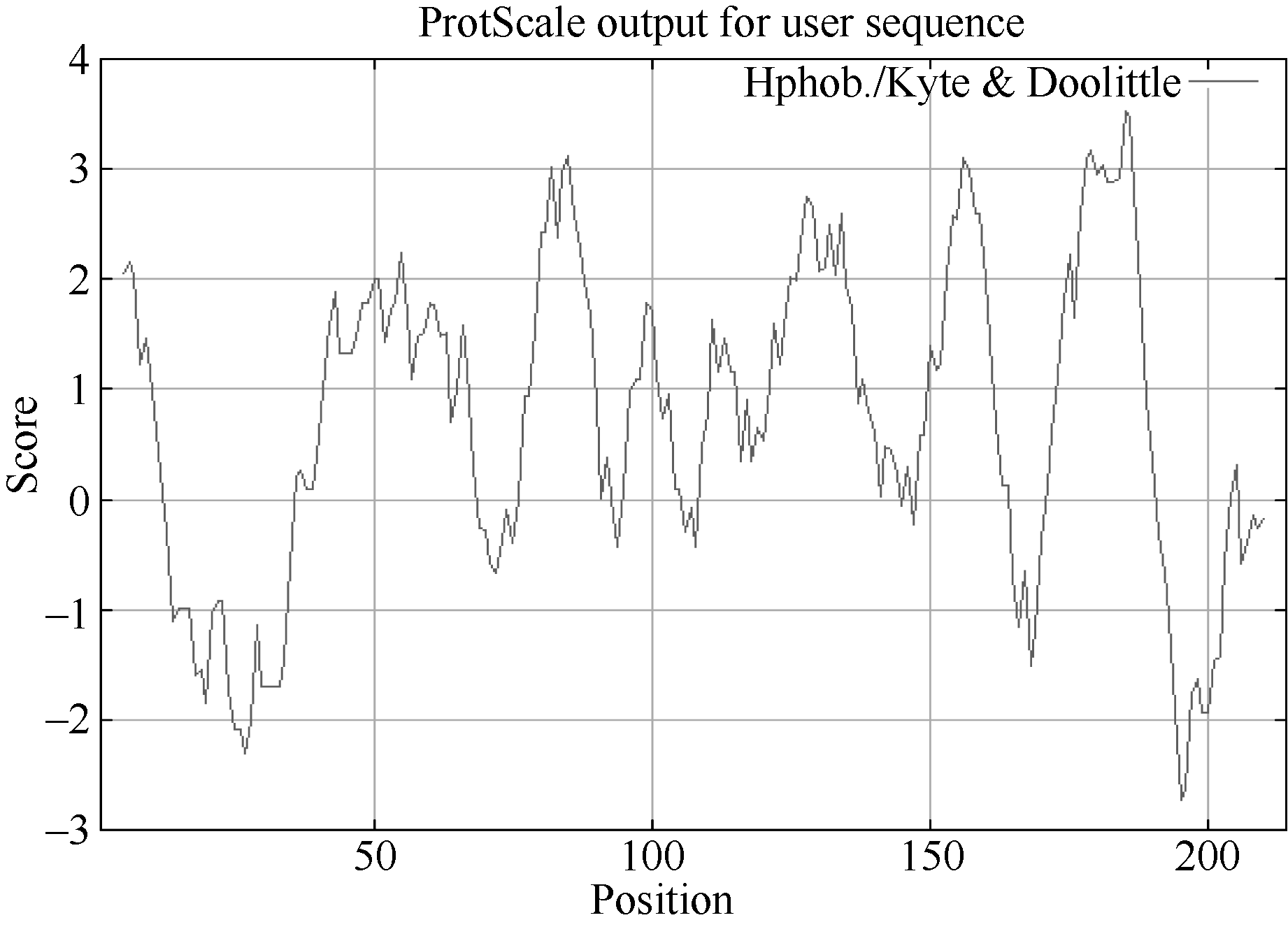
KyteJ, Doolittle RF. A simple method for displaying the hydropathic character of a protein. J Mol Biol, 1982, 157: 105-132[本文引用:1]
[40]
Nicoll DA, LongoniS, Philipson KD. Molecular cloning and functional expression of the cardiac sarcolemmal Na+-Ca2+ exchanger. Science, 1990, 250: 562-565[本文引用:1]
[41]
BlackfordS, Rea PA, Sand ersD. Voltage sensitivity of H+/Ca2+ antiport in higher plant tonoplast suggests a role in vacuolar calcium accumulation. J Biol Chem, 1990, 265: 9617-9620[本文引用:1]
[42]
Ettinger WF, Clear AM, Fanning KJ, Peck ML. Identification of a Ca2+/H+ antiport in the plant chloroplast thylakoid membrane. Plant Physiol, 1999, 119: 1379-1385[本文引用:1]
[43]
LiuH, Zhang XX, TakanoT, Liu SK. Characterization of a PutCAX1 gene from Puccinellia tenuiflora that confers Ca2+ and Ba2+ tolerance in yeast. Biochem Biophys Res Commun, 2009, 383: 392-396[本文引用:1]
[44]
XuL, Zahid KR, He LR, Zhang WW, HeX, Zhang XL, Yang XY, Zhu LF. GhCAX3 gene, a novel Ca2+/H+ exchanger from cotton, confers regulation of cold response and ABA induced signal transduction. PloS One, 2013, 8: e66303[本文引用:1]
[45]
Cheng NH, Pittman JK, ShigakiT, LachmansinghJ, LeClere S, Lahner B, Salt D E, Hirschi K D. Functional association of Arabidopsis CAX1 and CAX3 is required for normal growth and ion homeostasis. Plant Physiol, 2005, 138: 2048-2060[本文引用:1]
李国婧, 周燮. 水杨酸与植物抗非生物胁迫. 植物学通报, 2001, 18: 295-302Li GJ, ZhouX. Salicylic acid and abiotic stress resistance in plants. Chin Bull Bot, 2001, 18: 295-302 (in Chinese with English abstract)[本文引用:2]
[48]
张占军. PEG-6000模拟干旱胁迫下秋地黄瓜萌芽期抗旱性评价. 甘肃农业科技, 2014, (5): 16-18Zhang ZJ. Evaluation of the drought resistance of autumn cucumber in germination stage under PEG-6000 simulated drought stress. Gansu Agric Sci Techn, 2014, (5): 16-18[本文引用:1]
[49]
张和臣, 尹伟伦, 夏新莉. 非生物逆境胁迫下植物钙信号转导的分子机制. 植物学通报, 2007, 24: 114-122Zhang HC, Yin WL, Xia XL. The mechanism of Ca2+ signal transduction under abiotic stresses in plants. Chin Bull Bot, 2007, 24: 114-122 (in Chinese with English abstract)[本文引用:1]
[50]
陈沁, 刘友良. 谷胱甘肽对盐胁迫大麦叶片活性氧清除系统的保护作用. 作物学报, 2000, 26: 365-371ChenQ, Liu YL. Effect of glutathion on active oxygen scavenging system in leaves of barley seedlings under salt stress. Acta Agron Sin, 2000, 26: 365-371 (in Chinese with English abstract)[本文引用:1]
 , 刘峰
, 刘峰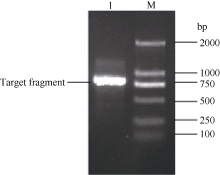








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}