关键词:小麦; 花发育; MADS-box基因; 表达模式 Expression Patterns of MADS-box Genes Related to Flower Development of Wheat LI Hai-Feng1,2,*, HAN Ying1, LIU Meng-Jia1, WANG Bing-Hua1, SU Ya-Li1, SUN Qi-Xin1,* 1 State Key Laboratory of Crop Stress Biology for Arid Areas / College of Agronomy, Northwest A&F University, Yangling 712100, China
2 Xinjiang Agriculture Vocational Technology College, Changji 831100, China
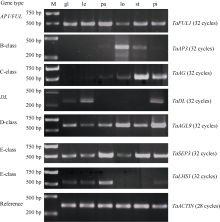
Fund:This study was supported by the National Natural Science Foundation of China (31571657), the Fundamental Research Foundation for the Central Universities (2014ZZ009), and the Foundation of Xinjiang Agriculture Vocational Technology College (XJNZYKJ201501) AbstractThe objective of this study was to elucidate the molecular mechanism of wheat ( Triticum aestivum L.) flower development. According to the phylogenetic tree of MADS- box genes from different species, we found that wheat contained all kinds of genes involved in the ABCDE model for flower development. The expression patterns of A-, B-, C-, D-, and E-class genes were analyzed by semi-quantitative and quantitative RT-PCR (qRT-PCR). Wheat AP1/ FUL gene TaFUL (A-class) was expressed in all floral organs with the highest expression level in lemmas and paleas. Genes TaAP3 (B-class), TaAG(C-class) showed conservative expression patterns in specific organs, i.e., TaAP3 was expressed in lodicules and stamens whereas TaAGwas expressed in stamens and pistils. The OsMADS13 homologous gene in wheat (D-class) was expressed in both pistils and lodicules, suggesting its function in lodicule and ovule development simultaneously. Gene TaSEP (E-class) was mainly expressed in paleas and the inner-three whorls. LHS1 is a grass-specific gene family and belongs to E-class. The expression of TaLHS1 was detected in lemmas, paleas, and glumes of wheat. TaDL, the homologous gene of rice DROOPING LEAF ( DL) controlling carpel development, was expressed in glumes, lemmas and carpels. These results suggest a conservative molecular mechanism for flower development in wheat, but some genes may have diversified functions due to evolution. The expression evidence of TaDL and TaLHS1 in glumes, in combination with the morphology and structure analyses of glume, lemma and palea, implied that lemma and glume might originate from the same organ in wheat.
图1 小麦花的结构和器官 A: 小穗; B: 剥去一半外稃的小花; C: 内轮花器官; D: 浆片; E: 护颖; F: 外稃; G: 內稃; H~J: 护颖(H)、外稃(I )和內稃(J)表层的扫描电镜照片; K: 小穗石蜡横切片; L和M: 护颖(L)和外稃(M)横切片局部放大; N: 內稃的横切片; O和P: 內稃边缘(O)和中间结构(P)的放大。L-N中, * 指示维管束。gl: 护颖; le: 外稃; pa: 內稃; lo: 浆片; st: 雄蕊; pi: 雌蕊。A~C和E~G中, bar = 500 µ m; D和H~J中, bar = 100 µ m。Fig. 1 Wheat flower structure and floral organs A: spikelet; B: floret with a half of lemma; C: inner floral organs; D: lodicules; E: glume; F: lemma; G: palea; H-J: epidermics SEM observation of glume (H), lemma (I), and palea (J); K: transverse section of one spikelet; L and M: Amplification of glume (L) and lemma (M). N: Transverse section of one palea; O and P: amplification of marginal tissue (O) and main strucuture of palea (P). Asterisks in L-N indicate vascular bundles. gl: glume; le: lemma; pa: palea; lo: lodicule; st: stamen; pi: pistil. Bar = 500 µ m in A-C and E-G; bar = 100 µ m in D and H-J.
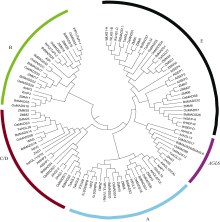
图2 拟南芥、水稻、短柄草、玉米、小麦花发育基因系统进化树 不同颜色的圆弧表示不同基因类型。Fig. 2 Phylogenetic tree of floral genes in Arabidopsis, Brachypodium distachyon, rice, maize, and wheat Color circular arcs show the gene classes.
Coen ES, Meyerowitz EM. The war of the whorls: genetic interactions controlling flower development. Nature, 199, 353: 31-37[本文引用:1]
[2]
PelazS, Ditta GS, BaumannE, WismanE, Yanofsky MF. B and C floral organ identity functions require SEPALLATA MADS-box genes. Nature, 2000, 405: 200-203[本文引用:2]
[3]
Theissen G. Development of floral organ identity: stories from the MADS house. Curr Opin Plant Biol, 2001, 4: 75-85[本文引用:2]
[4]
WangK, TangD, HongL, XuW, HuangJ, LiM, GuM, XueY, ChengZ. DEP and AFO regulate reproductive habit in rice. PLoS Genet, 2010, 6(1): e1000818[本文引用:1]
[5]
KobayashiK, YasunoN, SatoY, YodaM, YamazakiR, KimizuM, YoshidaH, NagamuraY, KyozukaJ. Inflorescence meristem identity in rice is specified by overlapping functions of three AP1/FUL-like MAD-box genes and PAP2, a SEPALLATA MADS-box gene. Plant Cell, 2012, 24: 1848-1859[本文引用:1]
[6]
NagasawaN, MiyoshiM, SanoY, SatohH, HiranoH, SakaiH, NagatoY. SUPERWOMAN1 and DROOPING LEAF genes control floral organ identity in rice. Development, 2003, 130: 705-718[本文引用:3]
[7]
YamaguchiT, LeeDY, MiyaoA, HirochikaH, AnG, Hirano HY. Functional diversification of the two C-class MADS-box genes OsMADS3 and OsMADS58 in Oryza sativa. Plant Cell, 2006, 18: 15-28[本文引用:1]
[8]
DreniL, JacchiaS, FornaraF, FornariM, Ouwerkerk PB, AnG, ColomboL, Kater MM. The D-lineage MADS-box gene OsMADS13 controls ovule identity in rice. Plant J, 2007, 52: 690-699[本文引用:1]
[9]
DreniL, PilatoneA, YunD, ErreniS, PajoroA, CaporaliE, ZhangD, Kater MM. Functional analysis of all AGAMOUS subfamily members in rice reveals their roles in reproductive organ identity determination and meristem determinacy. Plant Cell, 2011, 23: 2850-2863[本文引用:1]
[10]
LiH, LiangW, YinC, ZhuL, ZhangD. Genetic interaction of OsMADS3, DROOPING LEAF, and OsMADS13 in specifying rice floral organ identities and meristem determinacy. Plant Physiol, 2011, 156: 263-274[本文引用:4]
[11]
CuiR, HanJ, ZhaoS, SuK, WuF, DuX, XuQ, ChongK, TheissenG, MengZ. Functional conservation and diversification of class E floral homeotic genes in rice (Oryza sativa). Plant J, 2010, 61: 767-781[本文引用:3]
[12]
GaoX, LiangW, YinC, JiS, WangH, SuX, GuoC, KongH, XueH, ZhangD. The SEPALLATA-like gene OsMADS34 is required for rice inflorescence and spikelet development. Plant Physiol, 2010, 153: 728-740[本文引用:2]
[13]
LinX, WuF, DuX, ShiX, LiuY, LiuS, HuY, TheissenG, MengZ. The pleiotropic SEPALLATA-like gene OsMADS34 reveals that the ‘empty glumes’ of rice (Oryza sativa) spikelets are in fact rudimentary lemmas. New Phytol, 2014, 202: 689-702[本文引用:4]
[14]
YamaguchiT, NagasawaN, KawasakiS, MatsuokaM, NagatoY, Hirano HY. The YABBY gene DROOPING LEAF regulates carpel specification and midrib development in Oryza sativa. Plant Cell, 2004, 16: 500-509[本文引用:1]
[15]
LiH, LiangW, HuY, ZhuL, YinC, XuJ, DreniL, Kater MM, ZhangD. Rice MADS6 interacts with the floral homeotic genes SUPERWOMAN1, MADS3, MADS58, MADS13, and DROOPING LEAF in specifying floral organ identities and meristem fate. Plant Cell, 2011, 23: 2536-2552[本文引用:1]
[16]
王兆龙. 小麦小花发育的生理基础及调控研究. 南京农业大学博士学位论文, 江苏南京, 2000. pp 3-4Wang ZL. Physiological Basis and Regulation of Floret Development in Wheat. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2000. pp 3-4 (in Chinese with English abstract)[本文引用:1]
[17]
刘楠, 李海峰, 窦艳华, 韩德俊. 普通小麦及其近缘物种花序、小穗和小花的形态结构分析. 麦类作物学报, 2015, 35: 293-299LiuN, Li HF, Dou YH, Han DJ. Morphology and structure analyses on inflorescence, spikelet and floret of bread wheat and its relatives. J Triticeae Crops, 2015, 35: 293-299 (in Chinese with English abstract)[本文引用:1]
[18]
Paolacci AR, Tanzarella OA, PorcedduE, VarottoS, CiaffiM. Molecular and phylogenetic analysis of MADS-box genes of MIKC type and chromosome location of SEP-like genes in wheat (Triticum aestivum L. ). Mol Genet Genomics, 2007, 278: 689-708[本文引用:3]
[19]
IshikawaM, OhmoriY, TanakaW, HirabayashiC, MuraiK, OgiharaY, YamaguchiT, Hirano HY. The spatial expression patterns of DROOPING LEAF orthologs suggest a conserved function in grasses. Genes Genet Syst, 2009, 84: 137-146[本文引用:1]
[20]
ShitsukawaN, TahiraC, KassaiK, HirabayashiC, ShimizuT, TakumiS, MochidaK, KawauraK, OgiharaY, MuraiK. Genetic and epigenetic alteration among three homoeologous genes of a class E MADS-box gene in hexaploid wheat. Plant Cell, 2007, 19: 1723-1737[本文引用:5]
[21]
ZhaoT, NiZ, DaiY, YaoY, NieX, SunQ. Characterization and expression of 42 MADS-box genes in wheat (Triticum aestivum L. ). Mol Genet Genomics, 2006, 276: 334-350[本文引用:1]
[22]
Zhao XY, Cheng ZJ, Zhang XS. Overexpression of TaMADS1, a SEPALLATA-like gene in wheat, causes early flowering and the abnormal development of floral organs in Arabidopsis. Planta, 2006, 223: 698-707[本文引用:1]
[23]
TamuraK, PetersonD, PetersonN, StecherG, NeiM, KumarS. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol Biol Evol, 2011, 28: 2731-2739[本文引用:1]
[24]
窦艳华, 韩萌萌, 孙其信, 李海峰. 二穗短柄草MADS-BOX基因AGL6和FUL1的可变拼接和表达模式分析. 农业生物技术学报, 2015, 23: 459-468Dou YH, Han MM, Sun QX, Li HF. Alternative splicing and expression pattern analyses of two MADS-BOX genes AGL6 and FUL1 in Brachypodium distachyon. J Agric Biotechnol, 2015, 23: 459-468 (in Chinese with English abstract)[本文引用:3]
[25]
Li HF, Liang WQ, Jia RD, Yin CS, ZongJ, Kong HZ, Zhang DB. The AGL6-like gene OsMADS6 regulates floral organ and meristem identities in rice. Cell Res, 2010, 20: 299-313[本文引用:2]
[26]
JackT, Brockman LL, Meyerowitz EM. The homeotic gene APETALA3 of Arabidopsis thaliana encodes a MADS-box and is expressed in petals and stamens. Cell, 1992, 68: 683-697[本文引用:1]
[27]
Ambrose BA, Lerner DR, CiceriP, Padilla CM, Yanofsky MF, Schmidt RJ. Molecular and genetic analyses of the silky1 gene reveal conservation in floral organ specification between eudicots and monocots. Mol Cell, 2000, 5: 569-579[本文引用:1]
 , 韩英
, 韩英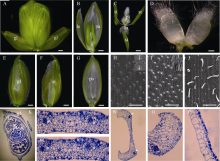






{kind=link}
{kind=link}
{kind=link}
{kind=link}