关键词:小麦; 近等基因系; 蜡质含量; 抗旱性 Drought Resistance of Wheat NILs with Different Cuticular Wax Contents in Flag Leaf XU Wen1,2, SHEN Hao1,2, GUO Jun2, YU Xiao-Cong2, LI Xiang1, YANG Yan-Hui1, MA Xiao1, ZHAO Shi-Jie1,*, SONG Jian-Min2,* 1State Key Laboratory of Crop Biology / College of Life Sciences, Shandong Agricultural University, Tai’an 271018, China
2 Crop Research Institute, Shandong Academy of Agricultural Sciences, Jinan 250100, China
Fund:This study was supported by the National Natural Science Foundation of China (31271635), the China Agriculture Research System (CARS-03-1-8), and the Innovation Project for Key Technologies in Shandong Province (2014GJJS0201-1) AbstractThe relationship of flag leaf cuticular wax content with leaf water potential, excised-leaf water loss, leaf photosynthesis characteristics and wheat yield under drought stress were analyzed in 2013-2014 and 2014-2015 wheat seasons using four wheat nearly isogenetic lines (NILs) differing in cuticular wax content in flag leaf. Under drought stress condition, the average flag leaf cuticular wax content of the high-wax NILs (15.15 mg g-1) was 79.8% higher than that of the low-wax NILs (8.43 mg g-1). Compared with the low-wax NILs, the high-wax NILs showed significantly higher water potential in flag leaf and lower water loss rate ( P< 0.05). In response to drought stress, photosynthetic rate ( Pn) and Fv/ Fm of flag leaf declined in both types of NILs, however, the decrease percentages were lower in the high-wax NILs (7.5% and 3.4%, respectively) than in the low-wax NILs (9.8% and 5.8%, respectively). As a result, the average yield of the high-wax NILs was 3.7% higher than that of the low-wax NILs, and the drought tolerance (TOL) and stress susceptible index (SSI) of the high-wax NILs were significantly less than those of the low-wax NILs ( P< 0.05). These results indicate that cuticular wax content in flag leaf is closely related to drought tolerance and can be used as a physiological indicator of drought resistance in wheat selection.
Keyword:Common wheat ( Triticum aestivum L.); Nearly isogenetic lines (NILs); Cuticular wax content; Drought resistance Show Figures Show Figures
图1 不同小麦近等基因系的表型和两种水分条件下旗叶蜡质含量比较 A图和B图为4个近等基因系开花期的田间表型, 显示叶片表面蜡质较少的JM204和JM206植株青绿, 颜色深于叶片表面蜡质较多的JM205和JM208。C图显示4个近等基因系在正常浇水和干旱胁迫下的叶片表面蜡质含量, 横坐标轴的1~4分别代表JM204、JM206、JM205和JM208, 误差线表示重复间的标准误差, 误差线上字母不同表示在相同水分条件下品系间差异显著(P < 0.05)。Fig. 1 Phenotypes of different NILs at flowering stage and comparison of flag leaf cuticular wax contents under two soil moisture conditions Panels A and B are the field phenotypes of four NILs at anthesis stage, showing JM204 and JM206 plants (lower cuticular wax content) are darker green than JM205 and JM208 plants (higher cuticular wax content). Panel C shows the difference of cuticular wax content in the four NILs under well-watered and drought stress conditions. The numbers 1 to 4 on X-axis represent lines JM204, JM206, JM205, and JM208, respectively. Error bars show standard error among three repeats, and different letters above error bars indicate significant difference among lines at P< 0.05.
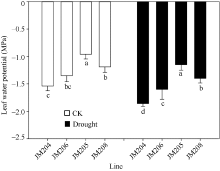
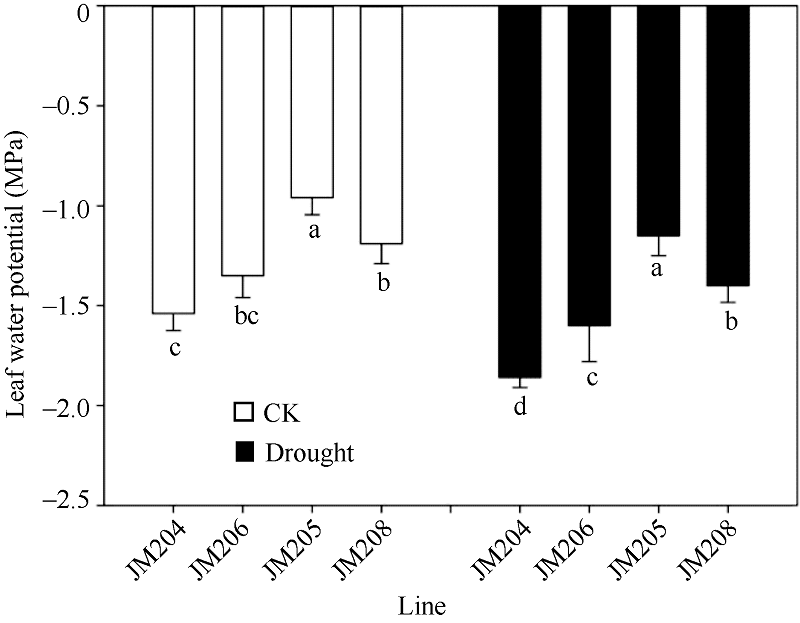
图2 正常浇水(CK)和干旱胁迫(drought)下不同小麦近等基因系灌浆中期旗叶水势比较 JM205和JM208为多蜡质品系, JM204和JM206为少蜡质品系。误差线表示重复间的误差, 误差线下字母不同表示品系间差异显著(P< 0.05)。Fig. 2 Comparison of flag leaf water potential of wheat NILs under well-watered (CK) and drought conditions in the middle stage of grain filling Different letters below the error bars indicate significant difference among lines (P< 0.05).
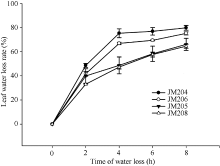
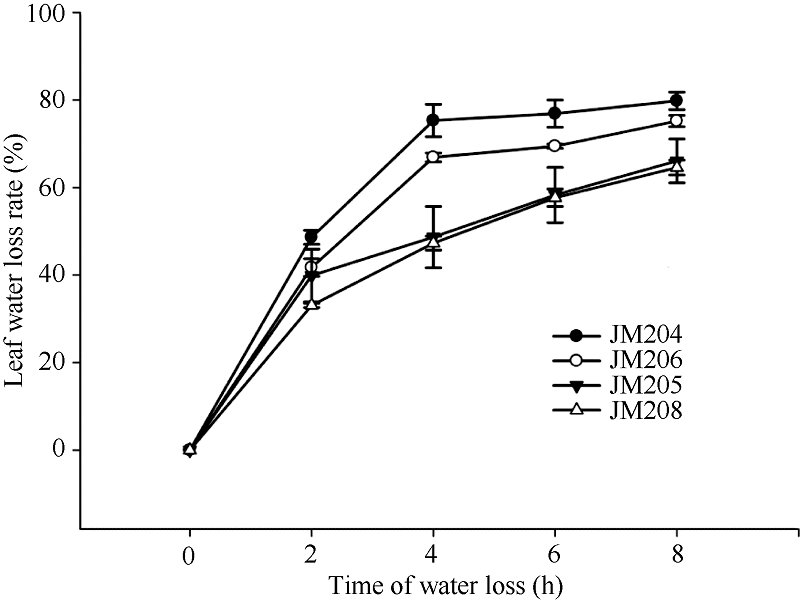
图3 干旱胁迫对不同小麦近等基因系灌浆中期旗叶水分散失速率的影响 JM205和JM208为多蜡质品系; JM204和JM206为少蜡质品系。Fig. 3 Effect of drought stress on water loss rate of flag leaf in wheat NILs in the middle stage of grain filling JM205 and JM208 are high-wax lines and JM204 and JM206 are low-wax lines.
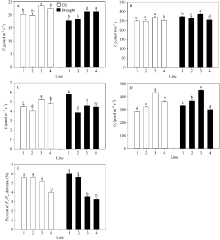
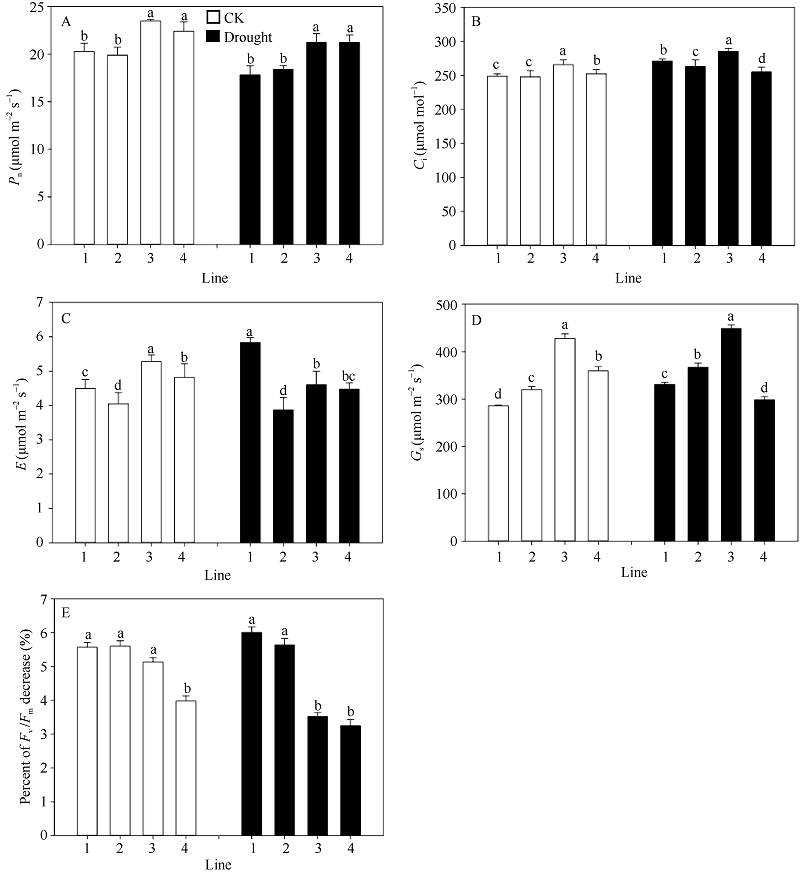
图4 不同蜡质含量小麦近等基因系旗叶光合速率Pn(A)、胞间CO2浓度Ci(B)、蒸腾速率E (C)、气孔导度Gs(D)和Fv/Fm(E) 横坐标轴的1~4分别表示品系JM204、JM206、JM205和JM208, 其中JM205和JM208为多蜡质品系, JM204和JM206为少蜡质品系。误差线表示重复间的标准误差, 误差线上字母不同表示品系间差异显著(P < 0.05)。Fig. 4 Flag leaf photosynthetic rate Pn (A), intercellular CO2 concentration Ci(B), transpiration rate E (C), stomatal conductance Gs(D), and Fv/Fm (E) in wheat NILs with different cuticular wax contents under well-watered and drought stress conditions The numbers 1 to 4 on X-axis represent lines JM204, JM206, JM205, and JM208, respectively. Error bars show standard error among three repeats, and different letters above error bars indicate significant difference among lines at P< 0.05.
表1 小麦不同蜡质含量近等基因系的籽粒产量和抗旱性 Table 1 Grain yield and drought tolerance of NILs with different cuticular wax contents
品系 Line
籽粒产量 Grain yield (kg hm-2)
抗旱指数 Tolerance
干旱敏感指数 Stress susceptible index
正常浇水 Well-watered
干旱胁迫 Drought
2013-2014
JM204
9096.21 b
8156.71 c
939.50 b
1.07 a
JM205
8854.43 c
8088.37 cd
766.06 c
0.90 b
JM206
9409.70 a
8221.71 b
1187.99 a
1.31 a
JM208
8932.80 c
8330.14 a
602.66 d
0.70 b
2014-2015
JM204
7856.98 a
6555.22 a
1301.76 a
1.25 b
JM205
7426.43 b
6540.77 a
885.66 b
0.90 c
JM206
6828.09 c
5480.40 b
1347.69 a
1.49 a
JM208
6614.20 cd
6355.96 a
258.24 c
0.30 d
JM205 and JM208 are high-wax lines and JM204 and JM206 are low-wax lines. In each growing season, values within a column followed by different letters are significantly different among lines at P< 0.05. JM205和JM208为多蜡质品系, JM204和JM206为少蜡质品系。同一年度中, 数据后字母不同表示品系间差异显著(P< 0.05)。
表1 小麦不同蜡质含量近等基因系的籽粒产量和抗旱性 Table 1 Grain yield and drought tolerance of NILs with different cuticular wax contents
图5 干旱胁迫条件下小麦少蜡质和多蜡质近等基因系灌浆后期田间生长情况 JM204 (A)和JM206 (B)为少蜡质品系; JM205 (C)和JM208 (D)为多蜡质品系。Fig. 5 Phenotype of wheat NILs with different cuticular wax contents in leaves at late grain-filling stage under drought stress JM204 (A) and JM206 (B) are low-wax lines; JM205 (C) and JM208 (D) are high-wax lines.
黄玲, 张正斌, 崔玉亭, 刘孟雨, 柴守玺, 陈兆波. 小麦叶片蜡质含量与水分利用效率和产量的关系. 麦类作物学报, 2003, 23(3): 41-44HuangL, Zhang ZB, Cui YT, Liu MY, Chai SX, Chen ZB. Relationship between cuticular wax content and water use efficiency of leaf and yield in wheat. J Triticeae Crops, 2003, 23(3): 41-44 (in Chinese with English abstract)[本文引用:2]
[2]
Johnson DA, Richards RA, Turner NC. Yield, water relations, gas exchange, and surface reflectances of near-isogenic wheat lines differing in glaucousness. Crop Sci, 1983, 23: 318-325[本文引用:1]
[3]
Foulkes MJ, Sylvester-BradleyR, WeightmanR, Weightman, Snape J W. Identifying physiological traits associated with improved drought resistance in winter wheat. Field Crops Res, 2007, 103: 11-24[本文引用:1]
柴凌燕, 董易凡, 李士伟, 史团省. 植物角质膜及其渗透性与抗旱性研究进展. 植物研究, 2010, 30: 763-768Chai LY, Dong YF, Li SW, Shi TX. Research progress in permeability and drought resistance of plant cuticular. Bull Bot Res, 2010, 30: 763-768 (in Chinese with English abstract)[本文引用:2]
[6]
李魏强, 张正斌, 李景娟. 植物表皮蜡质与抗旱及其分子生物学. 植物生理与分子生物学报, 2006, 32: 505-512Li WQ, Zhang ZB, Li JJ. Plant epicuticular cuticular wax and drought resistance as well as its molecular biology. J Plant Physiol Mol Biol, 2006, 32: 505-512 (in Chinese with English abstract)[本文引用:1]
[7]
李婧婧, 黄俊华, 谢树成. 植物蜡质及其与环境的关系. 生态学报, 2011, 31: 565-574Li JJ, Huang JH, Xie SC. Plant cuticular wax and its response to environmental conditions: an overview. Acta Ecol Sin, 2011, 31: 565-574 (in Chinese with English abstract)[本文引用:1]
[8]
PollardM, BeissonF, Li YH, Ohlrogge JB. Building lipid barriers: biosynthesis of cutin and suberin. Trends Plant Sci, 2008, 13: 236-246[本文引用:1]
[9]
KunstL, SamuelL. Plant cuticles shine: advances in cuticular wax biosynthesis and export. Plant Biol, 2009, 12: 721-727[本文引用:1]
[10]
BianchiG, LupottoE, BorghiB, CorbelliniM. Cuticular cuticular wax of wheat. The effects of chromosomal deficiencies on the biosynthesis of cuticular wax components. Planta, 1980, 148: 328-331[本文引用:1]
Premchand ra GS, SaneokaH, FujitaK, OgataS. Leaf water relations osmotic adjustment cell membrane stability epicuticular cuticular wax load and growth as affected by increasing water deficit in Sorghum. J Exp Bot, 1992, 43: 1569-1576[本文引用:1]
[13]
Bames JD, Percy KE, Poul ND. The influence of UV-B radiation on the physicochemical nature of tobacco (Nicotiana tabacum L. ) leaf surface. J Exp Bot, 1996, 47: 99-109[本文引用:1]
[14]
Eigenbrode SD, Espelie KE. Effects of plant epicuticular lipids on insect herbivores. Annu Rev Entomol, 1995, 40: 171-194[本文引用:3]
[15]
Kosma DK, JillA, Nemacheck. Changes in properties of wheat leaf cuticle during interactions with Hessian fly. Plant J, 2010, 63: 31-43[本文引用:3]
[16]
王美芳, 陈巨莲, 原国辉, 雷振生, 吴政卿, 赵献林. 植物表面蜡质对植食性昆虫的影响研究进展. 生态环境学报, 2009, 18: 1155-1160Wang MF, Chen JL, Yuan GH, Lei ZS, Wu ZQ, Zhao XL. Effects of plant epicuticular cuticular waxes on phytophagous insects behaviour. Ecol Environ Sci, 2009, 18: 1155-1160 (in Chinese with English abstract)[本文引用:3]
[17]
刘勇, 陈巨莲, 程登发. 不同小麦品种(系)叶片表面蜡质对两种麦蚜取食的影响. 应用生态学报, 2007, 18: 1785-1788LiuY, Chen JL, Cheng DF. Effects of wheat leaf surface cuticular waxes on the feeding of two wheat aphid species. Chin J Appl Ecol, 2007, 18: 1785-1788 (in Chinese with English abstract)[本文引用:3]
[18]
李希峰, 董娜. 小麦近等基因系的构建及应用进展. 安徽农业科学, 2012, 40: 2577-2579Li XF, DongN. Progress in construction and application of near-isogenic line in wheat. J Anhui Agric Sci, 2012, 40: 2577-2579 (in Chinese with English abstract)[本文引用:1]
[19]
Zadoks JC, Chang TT, Konzak CF. A decimal code for growth stages of cereals. Weed Res, 1974, 14: 415-421[本文引用:1]
[20]
Adamski NM, Bush MS, SimmondsJ, Turner AS, Mugford SG, JonesA, FindlayK, Pedentchouk N, von Wettstein-Knowles P, Uauy C. The Inhibitor of wax 1 locus (Iw1) prevents formation of β-and OH-β-diketones in wheat cuticular waxes and maps to a sub-cM interval on chromosome arm 2BS. Plant J, 2013, 746: 989-1002[本文引用:1]
[21]
ZhangZ, WangW, LiW. Genetic interactions underlying the biosynthesis and inhibition of β-diketones in wheat and their impact on glaucousness and cuticle permeability. PLoS One, 2013, 8: e54129[本文引用:2]
[22]
李鹏民, 高辉远, StrasserR. 快速叶绿素荧光诱导动力学分析在光合作用研究中的应用. 植物生理与分子生物学学报, 2005, 31: 559-566Li PM, Gao HY, StrasserR. Application of the fast chlorophyll fluorescence induction dynamics analysis in photosynthesis study. Acta Photophysiol Sin, 2005, 31: 559-566 (in Chinese with English abstract)[本文引用:1]
[23]
StrasserR, Tsimill-MichaelM, SrivastavaA. Analysis of the chlorophyll a fluorescence transient. In: Papageorgiou G C, Govindjee, eds. Advances in Photosynthesis and Respiration. Volume 19: Chlorophyll a Fluorescence: A Signature of Photosynthesis. Berlin: Springer, 2004. pp 321-362[本文引用:1]
[24]
Fischer RA, MaurerR. Drought resistance in spring wheat cultivars: I. Grain yield responses. Aust J Agric Res, 1978, 29: 897-912[本文引用:2]
[25]
Rosielle AA, HamblinJ. Theoretical aspects of selection for yield in stress and non-stress environment. Crop Sci, 1981, 21: 943-946[本文引用:1]
[26]
FleuryD, JefferiesS, KuchelH, LangridgeP. Genetic and genomic tools to improve drought tolerance in wheat. J Exp Bot, 2010, 61: 3211-3222[本文引用:2]
[27]
PiaoS, CiaisP, HuangY, HuangY, ShenZ, PengS, LiJ, ZhouL, LiuH, MaY, DingY, FriedlingsteinP, LiuC, TanK, YuY, ZhangT, FangJ. The impacts of climate change on water resources and agriculture in China. Nature, 2010, 467: 43-51[本文引用:2]
[28]
ShinozakiK, Yamaguchi-ShinozakiK, SekiM. Regulatory network of gene expression in the drought and cold stress responses. Curr Opin Plant Biol, 2003, 6: 410-417[本文引用:2]
[29]
Reddy AR, Chaitanya KV, Vivekanand anM. Drought-induced responses of photosynthesis and antioxidant metabolism in higher plants. J Plant Physiol, 2004, 161: 1189-1202[本文引用:1]
[30]
YordanovI, VelikovaV, TsonevT. Plant responses to drought, acclimation, and stress tolerance. Photosynthetica, 2000, 38: 171-186[本文引用:1]
[31]
ParentB, ShahinniaF, MaphosaL, BergerB, RabieH, ChalmersK, Langridge, FleuryD. Combining field performance with controlled environment plant imaging to identify the genetic control of growth and transpiration underlying yield response to water- deficit stress in wheat. J Exp Bot, 2015, 66: 5481-5492[本文引用:2]
[32]
高杨. 小麦叶片表皮蜡质的测定及其对光合和农艺性状的效应分析. 西北农林科技大学硕士论文, 陕西杨凌, 2014. pp 21-34GaoY. Determination of Epidermal Wax Content in Wheat Leaves and Analysis on Its Effect on Photosynthetic and Agronomic Traits. MS Thesis of Northwest A&F University, Yangling, China, 2014. pp 21-34 (in Chinese with English abstract)[本文引用:1]
[33]
Mir RR, Zaman-AllahM, SreenivasuluN, TrethowanR, Varshney RK. Integrated genomics, physiology and breeding approaches for improving drought tolerance in crops. Theor Appl Genet, 2012, 125: 625-645[本文引用:1]
[34]
TuberosaR, MaccaferriM. Genomics approaches to dissect the genetic basis of drought resistance in durum wheat. In: Yasunari Q, Shigeo T, Hirokazu H, eds. Advances in Wheat Genetics: from Genome to Field. Springer, Japan, 2015. pp 213-223[本文引用:1]
 , 宋健民
, 宋健民





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}