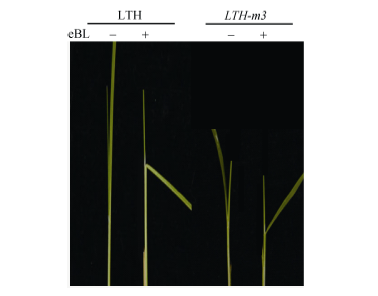
关键词:水稻; 突变体; 矮秆基因; 基因克隆; 基因功能 Genetic Identification of a New D1-allelic Mutant and Analysis of Its Gene Function in Rice WANG Cui-Hong, MA Jian, WANG Shuai, TIAN Peng, QI Chang-Yan, ZHAO Zhi-Chao, WANG Jiu-Lin, WANG Jie, CHENG Zhi-Jun, ZHANG Xin, GUO Xiu-Ping, LEI Cai-Lin* National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:This study was supported by the Special Fund for Agro-scientific Research in the Public Interest Program of China (201203014) and the Agricultural Science and Technology Innovation Program of Chinese Academy of Agricultural Sciences “Crop Functional Genomics” AbstractPlant height is one of important traits for rice yield. One genetically stable rice mutant, LTH-m3, was isolated from the cv. Lijiangxintuanheigu (LTH)-derived mutant population by mutagenesis using ethylmethane sulfonate (EMS). LTH-m3 was involved in the pathways of gibberellic acid (GA) and brassinosteroid (BR), and showed no sensitiveness to exogenous GA (GA3) and significantly reduced sensitiveness to exogenous BR (eBL) compared with the wild type. The genetic analysis, gene cloning and transgenic complementary test confirmed that LTH-m3 was a new d1-allelic mutant with small grain and dwarf phenotypes, and a single base was mutated (G2522→A2522) in the functional dwarf gene D1 at the conjunction site of its sixth exon and intron, which caused excision of the sixth exon in mRNA and premature termination of the D1encoded Gα protein, resulting in mutated phenotypes in the mutant. The further study showed that the D1 mutation caused obvious expression change of some dwarf genes such as SD1 and SLR1 in the mutant, and could affect the GA and BR pathways in their feedback regulations and signaling transductions in plant cells. The mutant overcome the defects of the universally blast-susceptible cv. LTH, such as too tall plant, soft stem and easy lodging, and could be utilized as an improved substitute of LTH in the future rice blast researches. The mutated D1 gene identified from the LTH-m3mutant may be useful for further study of Gα functions and signaling pathways of GA and BR.
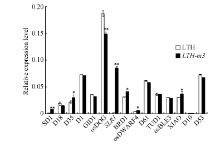
In the primer of SD1-Ex, “ SD1” stands for target gene, “ Ex” stands for the usage of the primer, i.e., gene expression, and the original name for the primer reported by Zhou et al. (2013)[46] is put in parenthesis. #: SD1-Ex中“ SD1” 代表目标基因, “ Ex” 代表引物的用途, 即用于基因表达分析, 括号内为Zhou等(2013)报道的引物原名[46]。
图1 野生型与突变体LTH-m3的表型 a, b: 幼苗; c: 第4叶片; d: 暗培养下形态; e: 成熟期植株; f: 穗; g: 粒宽; h: 粒长. 图a~f的左侧为野生型LTH, 右侧为突变体LTH-m3。图g和h的上侧为LTH, 下侧为LTH-m3。白色箭头指向中胚轴。Fig. 1 Phenotypes of the wild type and the LTH-m3 mutant a, b: seedlings; c: the 4th leaf; d: plant under dark treatment; e: adult plant; f: panicle; g: grain width; h: grain length. From figure a to f, the left is wild type LTH and the right is mutant LTH-m3; in figures g and h, the upper is wild type LTH and the lower is LTH-m3. The arrows indicate the mesocotyl.
表2 Table 2 表2(Table 2)
表2 野生型和LTH-m3的主要农艺性状比较 Table 2 Comparison of major agronomic traits between wild type and LTH-m3
性状 Trait
野生型 Wild type (LTH)
突变体 Mutant (LTH-m3)
抽穗期 Heading date (d)
94.0 ± 0.57
99.0 ± 0.67 * *
株高 Plant height (cm)
163.6 ± 3.50
57.7 ± 2.16 * *
分蘖数 Tillers per plant
18.8 ± 2.30
25.1 ± 4.23 * *
有效穗数 Panicles per plant
16.6 ± 2.17
19.5 ± 3.37 *
穗长 Panicle length (cm)
27.4 ± 6.40
19.2 ± 0.79 * *
穗粒数 Spikelets per panicle
161.6 ± 6.44
159.0 ± 10.79
结实率 Seed-setting rate (%)
92.2 ± 3.23
92.8 ± 4.43
粒长 Grain length (mm)
7.7 ± 0.28
5.0 ± 0.05 * *
粒宽 Grain width (mm)
3.3 ± 0.09
3.2 ± 0.02
千粒重 1000-grain weight (g)
23.0 ± 0.58
13.5 ± 0.26 * *
* : significantly different at P < 0.05, and * * : significantly different at P < 0.01 based on the t-test. * 表示t 测验在P < 0.05水平上差异显著, * * 表示t 测验在P< 0.01水平上差异显著。
表2 野生型和LTH-m3的主要农艺性状比较 Table 2 Comparison of major agronomic traits between wild type and LTH-m3
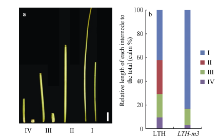
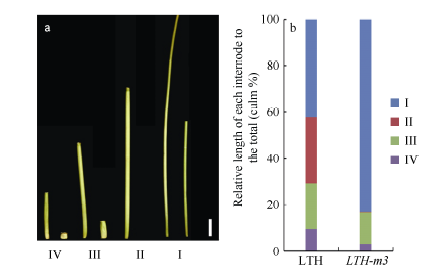
图2 野生型LTH和突变体LTH-m3各节间的长度及其伸长模式 a: LTH和LTH-m3各节间长度, 左侧为LTH, 右侧为LTH-m3, 标尺为5 cm。b: LTH和LTH-m3节间长度模式。I、II、III、IV分别表示穗下第1、第2、第3和第4节间。Fig. 2 Length and elongation model of the internodes in LTH and LTH-m3 a: internode length of LTH and LTH-m3. left: LTH; right: LTH-m3, the bar stands for 5 cm. b: internode elongation patterns of LTH and LTH-m3. I, II, III, and IV indicate internodes from the uppermost to the forth.
图3 GA3处理下突变体LTH-m3与野生型LTH苗高(a)及苗高增长率(b)的比较Fig. 3 Comparison of seedling height (a) and its relative increase (b) under treatments of GA3 between LTH-m3 mutant and wild type LTH
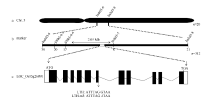
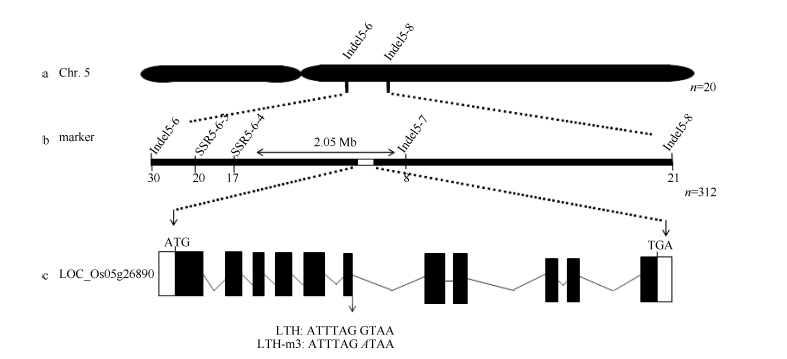
图5 突变体LTH-m3矮化基因的定位 a, b: 分别利用20个和312个突变表型的F2单株定位的基因位置。c: D1基因结构示意图, 白色框代表5′ 和3′ 非翻译区, 黑色框代表外显子区, 横线代表内含子区, 箭头指向突变体中D1的突变位点, 位于第6个内含子与外显子剪接位点处。Fig. 5 Mapping of the dwarf gene in mutant LTH-m3 a, b: gene mapping using 20 and 312 mutant-like F2 plants, respectively. c: the structure of D1, the white, black boxes represent 5′ and 3′ end un-translated regions and exons, and the horizontal lines represent introns. The arrow indicates the mutation position in the 6th intron of D1 in LTH-m3.
图7D1转基因互补表型 a: 抽穗期野生型、突变体及转基因阳性株T0代植株表型; b: 成熟期野生型、突变体及转基因阳性株T1代种子表型; c: 苗期野生型、突变体及转基因阳性株T1代植株表型。Fig. 7 Phenotypes complemented by transforming D1into the mutant a: phenotypes of the wild type, mutant and T0 transformant plants at heading stage; b: phenotypes of the wild type, mutant and T1 transformant seeds at maturing stage; c: phenotypes of the wild type, mutant and T0 transformant seedlings.
图6 突变体中D1基因的CDS序列和D1蛋白结构域突变位点 a: 野生型和突变体中D1基因CDS序列比对, 红色下画线部分为剪切掉的第6外显子。b: 突变体中D1蛋白结构域示意图, 第6外显子的缺失导致翻译提前终止, 箭头是翻译终止位置。Fig. 6 CDS sequence of D1 gene and the mutation site of D1 domain in the mutant a: alignment of CDS sequences of D1 in the mutant and wild type, in which the red underline indicates the cut 6th exon. b: a schematic diagram of the mutation in D1 protein, and the arrow indicates the mutation position of preterm stop codon in the mutant.
SasakiA, AshikariM, Ueguchi-TanakaM, ItohH, NishimuraA, SwapanD, IshiyamaK, SaitoT, KobayashiM, Khush GS, KitanoH, MatsuokaM. Green revolution: a mutant gibberellin-synthesis gene in rice. Nature, 2002, 416: 701-702[本文引用:1]
[2]
KhushG. Green revolution: Preparing for the 21st century. Genome, 1999, 42: 646-655[本文引用:1]
[3]
ZhangQ. Strategies for developing green super rice. Proc Natl Acad Sci USA, 2007, 104: 16402-16409[本文引用:1]
[4]
牛静, 陈赛华, 赵婕妤, 曾召琼, 蔡茂红, 周亮, 刘喜, 江玲, 万建民. 水稻株型突变体rad-1和rad-2的鉴定与功能基因克隆. 作物学报, 2015, 41: 1621-1631NiuJ, Chen SH, Zhao JY, Zeng ZQ, Cai MH, ZhouL, LiuX, JiangL, Wan JM. Identification and map-based cloning of rad-1 and rad-2, two rice architecture determinant mutants. Acta Agron Sin, 2015, 41: 1621-1631 (in Chinese with English abstract)[本文引用:2]
[5]
胡运高, 杨国涛, 郭连安, 钦鹏, 陈永军, 李世贵. 水稻多蘖突变体bf370的遗传分析和基因定位. 中国水稻科学, 2015, 29: 357-362Hu YG, Yang GT, Guo LA, QinP, Chen YJ, Li SG. Genetic analysis and mapping of a dwarf and high-tillering mutant bf370 in rice. Chin J Rice Sci, 2015, 29: 357-362 (in Chinese with English abstract)[本文引用:2]
[6]
NaganoH, OnishiK, OgasawaraM, HoriuchiY, SanoY. Genealogy of the “Green Revolution” gene in rice. Genes Genet Syst, 2005, 80: 1-6[本文引用:1]
[7]
罗茂春, 赵政, 夏令, 郭迟鸣, 陈亮. 水稻矮秆基因d-ss的遗传分析与克隆. 厦门大学学报(自然科学版), 2013, 52: 684-689Luo MC, ZhaoZ, XiaL, Guo CM, ChenL. Rice dwarfing genes and genetic analysis and cloning of d-ss. J Xiamen Univ (Nat Sci), 2013, 52: 684-689 (in Chinese with English abstract)[本文引用:3]
[8]
ChangT. Genetics and breeding. In: Westport. Rice: Production and Utilization. Connecticut: AVI Press, 1980. pp 146-187[本文引用:1]
ItohH, Ueguchi-TanakaM, SentokuN, KitanoH, MatsuokaM, KobayashiM. Cloning and functional analysis of two gibberellin 3β- hydroxylase genes that are differently expressed during the growth of rice. Proc Natl Acad Sci USA, 2001, 98: 8909-8914[本文引用:1]
GomiK, SasakiA, ItohH, Ueguchi-TanakaM, AshikariM, KitanoH, MatsuokaM. GID2, an F-box subunit of the SCF E3 complex, specifically interacts with phosphorylated SLR1 protein and regulates the gibberellin-dependent degradation of SLR1 in rice. Plant J, 2004, 37: 626-634[本文引用:1]
[21]
HiranoK, KouketuE, KatohH, AyaK, Ueguchi-TanakaM, MatsuokaM. The suppressive function of the rice DELLA protein SLR1 is dependent on its transcriptional activation activity. Plant J, 2012, 71: 443-453[本文引用:1]
[22]
ShimadaA, Ueguchi-TanakaM, SakamotoT. The rice spindly gene functions as a negative regulator of gibberellins signaling by controlling the suppressive function of the DELLA protein, SLR1, and modulating brassinosteroid synthesis. Plant J, 2006, 48: 390-402[本文引用:1]
[23]
TomonobuT, TakumaK, YukoH, MasamiU, ShihoA, KazunoriO, JinichiroK, WataruM, HisakazuY. Cloning and characterization of cDNAs encoding ent-copaly diphosphate synthases in wheat: insight into the evolution of rice phytoalexin biosynthetic genes. Biosci Biotechnol Biochem, 2009, 73: 772-775[本文引用:1]
[24]
Mallikarjuna RK, Zhang YS, Yu SB, Yan WH, Xing YZ. Cand idacy of a chitin-inducible gibberellin-responsive gene for a major locus affecting plant height in rice that is closely linked to green revolution gene sd1. Theor Appl Genet, 2011, 123: 705-714[本文引用:1]
[25]
Liu YJ, Xu YY, XiaoJ, Ma QB, LiD, XueZ, ChongK. OsDOG, a gibberellin-induced A20/AN1 zinc-finger protein, negatively regulates gibberellin-mediated cell elongation in rice. J Plant Physiol, 2011, 168: 1098-1105[本文引用:1]
[26]
HuangJ, TangD, ShenY, Qin BX, Hong LL, You AQ, LiM, WangX, Yu HX, Gu MH, Cheng ZK. Activation of gibberellins 2-oxidase 6 decreases active gibberellin levels and creates a dominant semi-dwarf phenotype in rice (Oryza sativa L. ). J Genet Genomics, 2010, 37: 23-26[本文引用:1]
[27]
MarciaM, ZhouX, ZhuQ, DennisE, UpadhyayaN. Isolation and characterization of a Ds-tagged rice (Oryza sativa L. ) GA-responsive dwarf mutant defective in an early step of the gibberellin biosynthesis pathway. Plant Cell Rep, 2005, 23: 819-833[本文引用:2]
[28]
Ueguchi-TanakaM, FujisawaY, KobayashiM, AshikariM, IwasakiY, KitanoH, MatsuokaM. Rice dwarf mutant d1, which is defective in the alpha subunit of the heterotrimeric G protein, affects gibberellin signal transduction. Proc Natl Acad Sci USA, 2000, 97: 11638-11643[本文引用:3]
[29]
HongZ, Ueguchi-TanakaM, UmemuraK, UozuS, FujiokaS, TakatsutoS, YoshidaS, AshikariM, KitanoH, MatsuokaM. A rice brassinosteroid-deficient mutant, ebisu dwarf (d2), is caused by a loss of function of a new member of cytochrome P450. Plant Cell, 2003, 15: 2900-2910[本文引用:4]
NakagawaH, AtsunoriT, TakanariT, OhtakeM, FujiokaS, NakamuraH, IchikawaH, MasakiM. Short grain1 decrease organ elongation and brassinosteroid response in rice. Plant Physiol, 2012, 158: 1208-1219[本文引用:1]
[32]
TongH, JinY, LiuW, LiF, FangJ, Yin YH, QianQ, Zhu LH, Chu CC. Dwarf and low-tillering, a new member of the GRAS family, plays positive roles in brassinosteroid signaling in rice. Plant J, 2009, 58: 803-816[本文引用:1]
[33]
BaiM, ZhangL, Srinivas SG, Zhu SW, Song WY, ChongK, WangZ. Functions of OsBZR1 and 14-3-3 proteins in brassinosteroid signaling in rice. Proc Natl Acad Sci USA, 2007, 104: 13839-3844[本文引用:1]
[34]
HuX, QianQ, XuT, ZhangE, Dong GJ, GaoT, QiX, Xue YB. The U-box E3 ubiquitin ligase TUD1 funcations with a heterotrimeric Gα-subunit to brassinosteroid-mediated growth in rice. PLos Genet, 2013, 9(3): e1003391[本文引用:2]
[35]
Sui PF, JinJ, YeS, MuC, GaoJ, Feng HY, Shen WH, YuY, Dong AW. H3K36 methylation is critical for brassinosteroid- regulated plant growth and development in rice. Plant J, 2012, 70: 340-347[本文引用:1]
[36]
Jiang YH, BaoL, So-YoonJ, Seong-KiK, Xu CG, Li XH, Zhang QF. XIAO is involved in the control of organ size by contributing to the regulation of signaling and homeostasis of brassinosteroids and cell cycling in rice. Plant J, 2012, 70: 398-408[本文引用:2]
[37]
YangG, NakamuraH, IchikawaH, KitanoH, KomatsuS. OsBLE3, a brassinolide-enhanced gene, is involved in the growth of rice. Phytochemistry, 2006, 67: 1442-1454[本文引用:1]
[38]
TanabeS, AshikariM, FujiokaS, TakatsutoS, YoshidaS, YanoM, YoshimuraA, KitanoH, MatsuokaM, FujisawaY, KatoH, IwasakiY. A novel cytochrome P450 is implicated in brassinosteroid biosynthesis via the characterization of a rice dwarf mutant, dwarf11, with reduced seed length. Plant Cell, 2005, 17: 776-790[本文引用:1]
[39]
AriteT, UmeharaM, IshikawaS, HanadaA, MaekawaM, YamaguchiS, Kyozuka J. d14, a strigolactone insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol, 2009, 50: 1416-1424[本文引用:1]
[40]
YamamuroC, IharaY, WuX, NoguchiT, FujiokaS, TakatsutoS, AshikariM, KitanoH, MatsuokaM. Loss of function of a rice brassinosteroid insensitive1 homolog prevents internode elongation and bending of the lamina joint. Plant Cell, 2000, 12: 1591-1606[本文引用:2]
[41]
SakamotoT, MorinakaY, InukaiY, KitanoH, FujiokaS. Auxin signal transcription factor regulates expression of the brassinosteroid receptor gene in rice. Plant J, 2013, 73: 676-688[本文引用:2]
ZouJ, ZhangS, ZhangW, LiG, ChenZ, ZhaiW, ZhaoX, PanX, XieQ, ZhuL. The rice HIGH-TILLERING DWARF1 encoding an ortholog of Arabidopsis MAX3 is required for negative regulation of the outgrowth of axillary buds. Plant J, 2006, 48: 687-698[本文引用:1]
[44]
LinH, WangR, QianQ, YanM, MengX, FuZ, YanC, JiangB, SuZ, LiJ, WangY. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell, 2009, 21: 1512-1525[本文引用:1]
[45]
Guo SY, Xu YY, Liu HH, Mao ZW, ZhangC, MaY, Zhang QR, MengZ, ChongK. The interaction between OsMADS57 and OsTB1 modulates rice tillering via DWARF14. Nat Commun, 2013, 4: 1566[本文引用:1]
OkiK, InabaN, KitagawaK, FujiokaS, KitanoH, FujisawaY, KatoH, IwasakiY. Function of the α subunit of rice heterotrimeric G protein in Brassinosteroid signaling. Plant Cell Physiol, 2009, 50: 161-172[本文引用:6]
[48]
AshikariM, Wu JZ, YanoM, SasakiT, YoshimuraA. Rice gibberellin-insensitive dwarf mutant gene Dwarf1 encodes the α-subunit of GTP-binding protein. Proc Natl Acad Sci USA, 1999, 96: 10284-10289[本文引用:2]
[49]
FujisawaY, KatoT, OhkiS, IshikawaA, KitanoH, SasakiT, AsahiT, IwasakiY. Suppression of the heterotrimeric G protein causes abnormal morphology, including dwarfism, in rice. Proc Natl Acad Sci USA, 1999, 96: 7575-7580[本文引用:3]
[50]
AssmannS M. G protein regulation of disease resistance during infection of rice with rice blast fungus. Sci STKE, 2005, 310: cm13[本文引用:2]
[51]
Jones AM, Assmann SM. Plants: the latest model system for G-protein research. EMBO Rep, 2004, 5: 572-578[本文引用:2]
[52]
Assmann SM. G proteins go green: a plant G protein signaling FAQ sheet. Science, 2005, 310: 71-73[本文引用:2]
[53]
SuharsonoU, FujisawaY, KawasakiT, IwasakiY, SatohH, ShimamotoK. The heterotrimeric G protein α subunit acts upstream of the small GTPase Rac in disease resistance of rice. Proc Natl Acad Sci USA, 2002, 99: 13307-13312[本文引用:1]
[54]
LieberherrD, ThaoN, NakashimaA, UmemuraK, KawasakiT, ShimamotoK. A sphingolipid elicitor-inducible mitogen- activated protein kinase is regulated by the small GTPase OsRac1 and heterotrimeric G-protein in rice. Plant Physiol, 2005, 138: 1644-1652[本文引用:1]
[55]
IwasakiY, FujisawaY, KatoH. Function of heterotrimeric G protein in gibberellin signaling. Plant Growth Regul, 2003, 22: 126-133[本文引用:1]
[56]
LingZ, MewT, WangJ, LeiC, HangN. Development of Chinese near-isogenic lines of rice and their differentiating ability of pathogenic races of blast fungus. Chin Agric Sci, 2001, 1: 50-56[本文引用:2]
[57]
KobayashiN, Telebanco-Yanoria M J, Tsunematsu H, Kato H, Imbe T, Fukuta Y. Development of new sets of international stand ard differential varieties for blast resistance in rice (Oryza sativa L. ). Jpn Agric Res Q, 2007, 41: 31-37[本文引用:2]
[58]
OkiK, InabaN, KitagawaK, KitanoH, TakahashiS, FujisawaY, KatoH, IwasakiY. Study of novel d1 alleles, defective mutants of the α subunit of heterotrimeric G-protein in rice. Genes Genet Syst, 2009, 84: 35-42[本文引用:2]
[59]
纪现军, 叶胜海, 周涯, 修芬连, 邓晓梅, 尚海漩, 刘继云, 陈萍萍, 李小华, 金庆生, 张小明. 水稻矮秆突变体Zj88d的鉴定与基因定位. 中国水稻科学, 2013, 27: 35-40Ji XJ, Ye SH, ZhouY, Xiu FL, Deng XM, Shang HX, Liu JY, Chen PP, Li XH, Jin QS, Zhang XM. Characterization and gene mapping of a dwarf mutant Zj88d in rice. Chin J Rice Sci, 2013, 27: 35-40 (in Chinese with English abstract)[本文引用:1]
[60]
侯雷, 袁守江, 尹亮, 赵金凤, 万国峰, 张文会, 李学勇. 两个新水稻Dwarf18基因强等位突变体的表型分析及分子鉴定. 作物学报, 2013, 38: 1416-1424HouL, Yuan SJ, YinL, Zhao JF, Wan GF, Zhang WH, Li XY. Phenotypic analysis and molecular characterization of two allelic mutants of the dwarf18 gene in rice. Acta Agron Sin, 2013, 38: 1416-1424 (in Chinese with English abstract)[本文引用:1]
[61]
马良勇, 包劲松, 李西明, 朱旭东, 季芝娟, 夏英武, 杨长登. 水稻矮生基因的克隆和功能研究进展. 中国水稻科学, 2009, 23: 1-11Ma LY, Bao JS, Li XM, Zhu XD, Ji ZJ, Xia YW, Yang CD. Progress on cloning and functional analysis of dwarfism related genes in rice. Chin J Rice Sci, 2009, 23: 1-11 (in Chinese with English abstract)[本文引用:3]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}