关键词:小麦; 蛋白磷酸酶2A; 启动子; 顺式作用元件; GUS组织化学染色; GUS定量分析 Cloning and Expression Analysis of Protein Phosphatase 2A Gene TaPP2AbB″-α Promoter in Wheat YI Heng1,2, LI Ang2, LIU Hui-Min1, JING Rui-Lian2,* 1 College of Bioengineering, Shanxi University, Taiyuan 030006, China
2 National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
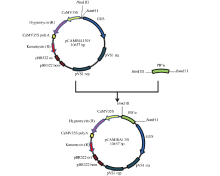
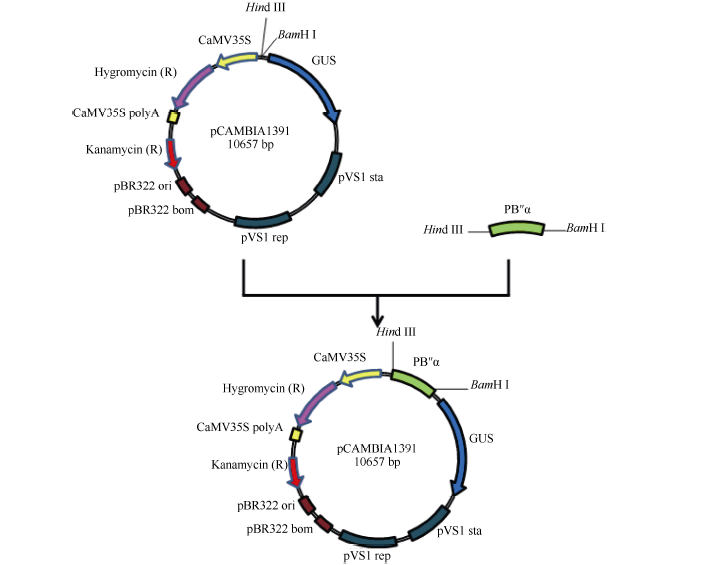
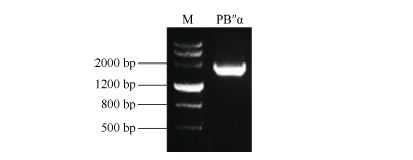
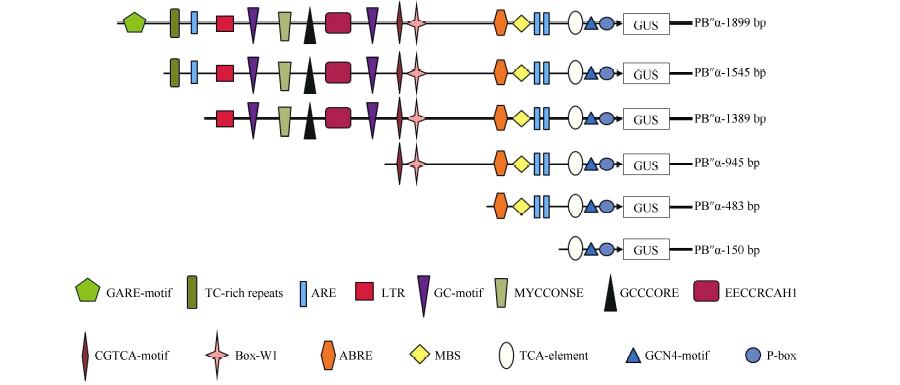
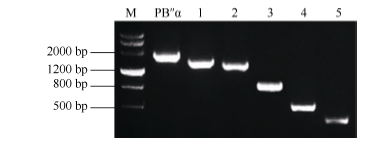
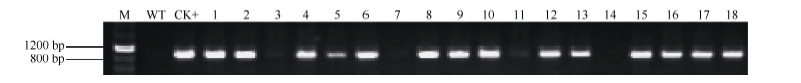
Fund:This study was supported by the National Natural Science Foundation of China (31201206) and the Agricultural Science and Technology Innovation Program (CAAS) AbstractProtein phosphatase 2A (PP2A) is a heterotrimeric protein, consisting of a scaffolding subunit (A), a catalytic subunit (C), and a member of four families of regulatory subunits (B). PP2A plays significant roles in the pathway responding to abiotic stresses in plants. TaPP2AbB″-α, a member of regulatory subunit B″ in wheat ( Triticum aestivum L.), enhanced root development and could develop more lateral roots in the gene overexpressed Arabidopsis, especially under the osmotic stresses of mannitol and NaCl. In order to elucidate transcriptional regulatory mechanism of the promoter PB″α of TaPP2AbB″-α, we isolated an 1899 bp full-length sequence of promoter PB″α from a drought-tolerant wheat cultivar Hanxuan 10. The PB″α sequence contained TATA-box, CAAT-box and a series of cis-acting elements responding to drought and osmotic stresses, such as elements of EECCRCAH1 (from -1058 to -1052 bp), GCCCORE (from -1073 to -1068 bp) and MYCCONSE (from -1179 to -1174 bp). The full-length promoter PB″α and five 5′-end truncated PB″α promoters in different lengths fused with the reporter gene β-glucuronidas ( GUS) were transformed into Arabidopsis, respectively. The histochemical staining results showed that the full-length promoter PB″α, and the deletion fragments of PB″α-1545 and PB″α-1389 could drive GUS gene expression in shoots and roots of transgenic Arabidopsis seedlings. As the result of quantitative fluorometric GUS assay, only the expression of PB″α, PB″α-1545 and PB″α-1389 could be up-regulated by salt and osmotic stresses in transgenic Arabidopsis lines, and the active region of PB″α promoter was located in the interval between -1389 bp and -946 bp. In conclusion, PB″α has strong basic promoter activity which is up-regulated significantly by salt and osmotic stresses. These findings contribute to the selection of a suitable promoter for crop improvement.
Keyword:Wheat; PP2A; Promoter; cis-acting Element; GUS histochemical staining; Quantitative fluorometric GUS assay Show Figures Show Figures
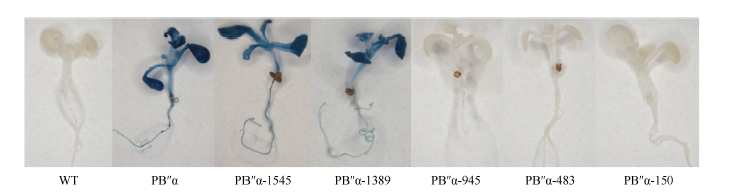
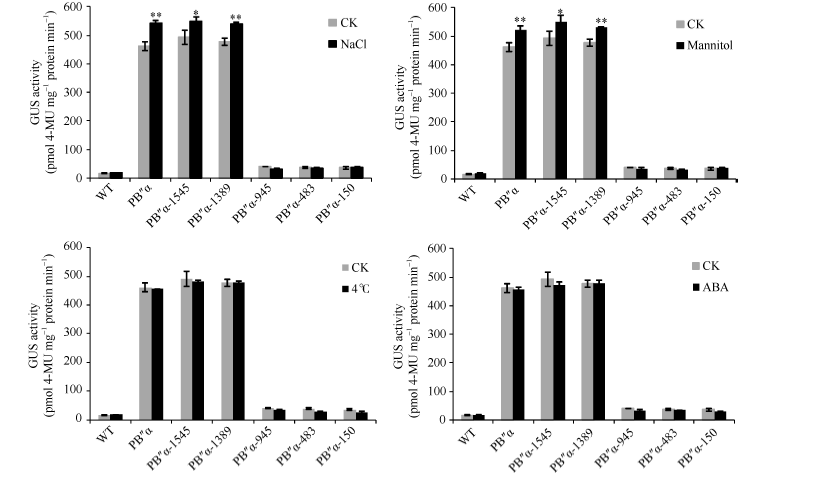
图8 胁迫条件下转基因拟南芥GUS定量分析 * 和* * 分别表示胁迫处理与对照(CK)在0.05和0.01概率水平的差异显著(t检验)。Fig. 8 Quantification of GUS activity in transgenic Arabidopsis under different stresses * and * * indicate significant difference between stress treatment and the control (CK) at the 0.05 and 0.01 probability levels, respectively (t-test).
ShinozakiK, Yamaguchi-ShinozakiK. Gene networks involved in drought stress response and tolerance. J Exp Bot, 2007, 58: 221-227[本文引用:1]
[2]
JanssensV, GorisJ. Protein phosphatase 2A: a highly regulated family of serine/threonine phosphatases implicated in cell growth and signalling. Biochem J, 2001, 353: 417-439[本文引用:1]
[3]
Høyer-HansenM, BastholmL, SzyniarowskiP, CampanellaM, SzabadkaiG, FarkasT, BianchiK, FehrenbacherN, EllingF, RizzutoR, Mathiasen IS, JäätteläM. Control of macroautophagy by calcium, calmodulin-dependent kinase kinase-beta, and Bcl-2. Mol Cell, 2007, 25: 193-205[本文引用:1]
[4]
BallesterosI, DominguezT, SauerM, ParedesP, DupratA, RojoE, SanmartinM, Sanchez-Serrano J J. Specialized functions of the PP2A subfamily II catalytic subunits PP2A-C3 and PP2A-C4 in the distribution of auxin fluxes and development in Arabidopsis. Plant J, 2013, 73: 862-872[本文引用:2]
[5]
TangW, YuanM, WangR, YangY, WangC, Oses-Prieto J A, Kim T W, Zhou H W, Deng Z, Gampala S S, Gendron J M, Jonassen E M, Lillo C, DeLong A, Burlingame A L, Sun Y, Wang Z Y. PP2A activates brassinosteroid-responsive gene expression and plant growth by dephosphorylating BZR1. Nat Cell Biol, 2011, 13: 124-131[本文引用:1]
Chavez-Aviles MN, Andrade-Perez CL, de la Cruz HR. PP2A mediates lateral root development under NaCl-induced osmotic stress throughout auxin redistribution in Arabidopsis thaliana. Plant & Soil, 2013, 368: 591-602[本文引用:1]
[8]
XuC, JingR, MaoX, JiaX, ChangX. A wheat (Triticum aestivum) protein phosphatase 2A catalytic subunit gene provides enhanced drought tolerance in tobacco. Ann Bot, 2007, 99: 439-450[本文引用:1]
[9]
ShiY. Serine/threonine phosphatases: mechanism through structure. Cell, 2009, 193: 468-484[本文引用:1]
[10]
Slupe AM, Merrill RA, StrackS. Determinants for substrate specificity of protein phosphatase 2A. Enzyme Res, 2011, 2011: 398751[本文引用:1]
[11]
Pand eyS, Mahato PK, BhattacharyyaS. Metabotropic glutamate receptor 1 recycles to the cell surface in protein phosphatase 2A-dependent manner in non-neuronal and neuronal cell lines. J Neurochem, 2014, 131: 602-614[本文引用:1]
[12]
Uhrig RG, Laband era AM, Moorhead GB. Arabidopsis PPP family of serine/threonine protein phosphatases: many targets but few engines. Trends Plant Sci, 2013, 18: 505-513[本文引用:1]
[13]
FarkasI, DombradiV, MiskeiM, SzabadosL, KonczC. Arabidopsis PPP family of serine/threonine phosphatases. Trends Plant Sci, 2007, 12: 169-176[本文引用:1]
[14]
Harris DM, Myrick TL, Rundle SJ. The Arabidopsis homolog of yeast TAP42 and mammalian alpha4 binds to the catalytic subunit of protein phosphatase 2A and is induced by chilling. Plant Physiol, 1999, 121: 609-617[本文引用:1]
[15]
LiuD, LiA, MaoX, JingR. Cloning and characterization of TaPP2AbB"-alpha, a member of the PP2A regulatory subunit in wheat. PLoS One, 2014, 9: e94430[本文引用:2]
[16]
Benfey PN, Chua NH. The Cauliflower mosaic virus 35S promoter: combinatorial regulation of transcription in plants. Science, 1990, 250: 959-966[本文引用:1]
[17]
Estrada-Melo AC, MaC, Reid MS, Jiang CZ. Overexpression of an ABA biosynthesis gene using a stress-inducible promoter enhances drought resistance in petunia. Hort Res, 2015, 2: 15013[本文引用:1]
[18]
NakashimaK, JanA, TodakaD, MaruyamaK, GotoS, ShinozakiK, Yamaguchi-ShinozakiK. Comparative functional analysis of six drought-responsive promoters in transgenic rice. Planta, 2014, 239: 47-60[本文引用:1]
[19]
LiM, WangX, CaoY, LiuX, LinY, OuY, ZhangH, LiuJ. Strength comparison between cold-inducible promoters of Arabidopsis cor15a and cor15b genes in potato and tobacco. Plant Physiol Biochem, 2013, 71: 77-86[本文引用:1]
[20]
Hernand ez-Garcia CM, Finer JJ. A novel cis-acting element in the GmERF3 promoter contributes to inducible gene expression in soybean and tobacco after wounding. Plant Cell Rep, 2015, 35: 1-14[本文引用:1]
[21]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4325[本文引用:1]
[22]
Sieburth LE, Meyerowitz EM. Molecular dissection of the AGAMOUS control region shows that cis elements for spatial regulation are located intragenically. Plant Cell, 1997, 9: 355-365[本文引用:1]
[23]
Vasil IK, VasilV. Transformation of wheat via particle bombardment. Methods Mol Biol, 2006, 318: 273-283[本文引用:1]
[24]
Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein dye binding. Anal Biochem, 1976, 72: 248-254[本文引用:1]
[25]
LuoK, ZhangG, DengW, LuoF, QiuK, PeiY. Functional characterization of a cotton late embryogenesis-abundant D113 gene promoter in transgenic tobacco. Plant Cell Rep, 2008, 27: 707-717[本文引用:1]
[26]
SinghH, SenR, BaltimoreD, Sharp PA. A nuclear factor that binds to a conserved sequence motif in transcriptional control elements of immunoglobulin genes. Nature, 1986, 319: 154-158[本文引用:1]
[27]
Li JJ, HerskowitzI. Isolation of ORC6, a component of the yeast origin recognition complex by a one-hybrid system. Science, 1993, 262: 1870-1874[本文引用:1]
[28]
ChakravarthyS, Tuori RP, D’Ascenzo M D, Fobert P R, Despres C, Martin G B. The tomato transcription factor Pti4 regulates defense-related gene expression via GCC box and non-GCC box cis elements. Plant Cell, 2003, 15: 3033-3050[本文引用:1]
[29]
AbeH, UraoT, ItoT, SekiM, ShinozakiK, Yamaguchi-ShinozakiK. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell, 2003, 15: 63-78[本文引用:1]
[30]
Kim HJ, Kim YK, Park JY, KimJ. Light signalling mediated by phytochrome plays an important role in cold-induced gene expression through the C-repeat/dehydration responsive element (C/DRE) in Arabidopsis thaliana. Plant J, 2002, 29: 693-704[本文引用:1]
[31]
BihmidineS, LinJ, Stone JM, AwadaT, Specht JE, Clemente TE. Activity of the Arabidopsis RD29A and RD29B promoter elements in soybean under water stress. Planta, 2013, 237: 55-64[本文引用:1]
[32]
KovalchukN, JiaW, EiniO, MorranS, PyvovarenkoT, FletcherS, BazanovaN, HarrisJ, Beck-OldachK, ShavrukovY, LangridgeP, LopatoS. Optimization of TaDREB3 gene expression in transgenic barley using cold-inducible promoters. Plant Biotechnol J, 2013, 11: 659-670[本文引用:1]
[33]
YiN, Oh SJ, Kim YS, Jang HJ, Park SH, Jeong JS, Song SI, Choi YD, Kim JK. Analysis of the Wsi18, a stress-inducible promoter that is active in the whole grain of transgenic rice. Transgenic Res, 2011, 20: 153-163[本文引用:1]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}