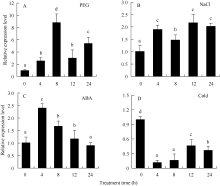
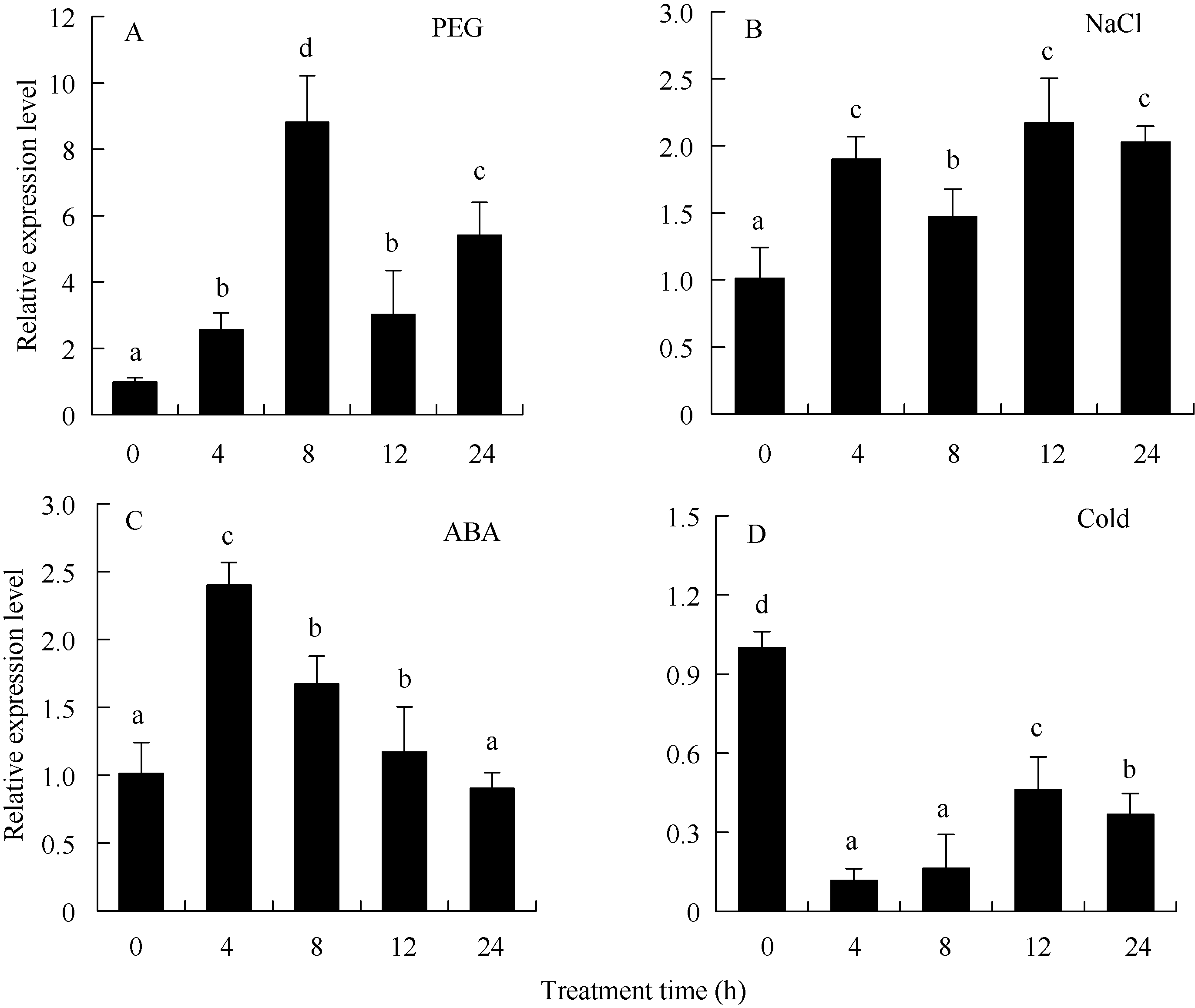
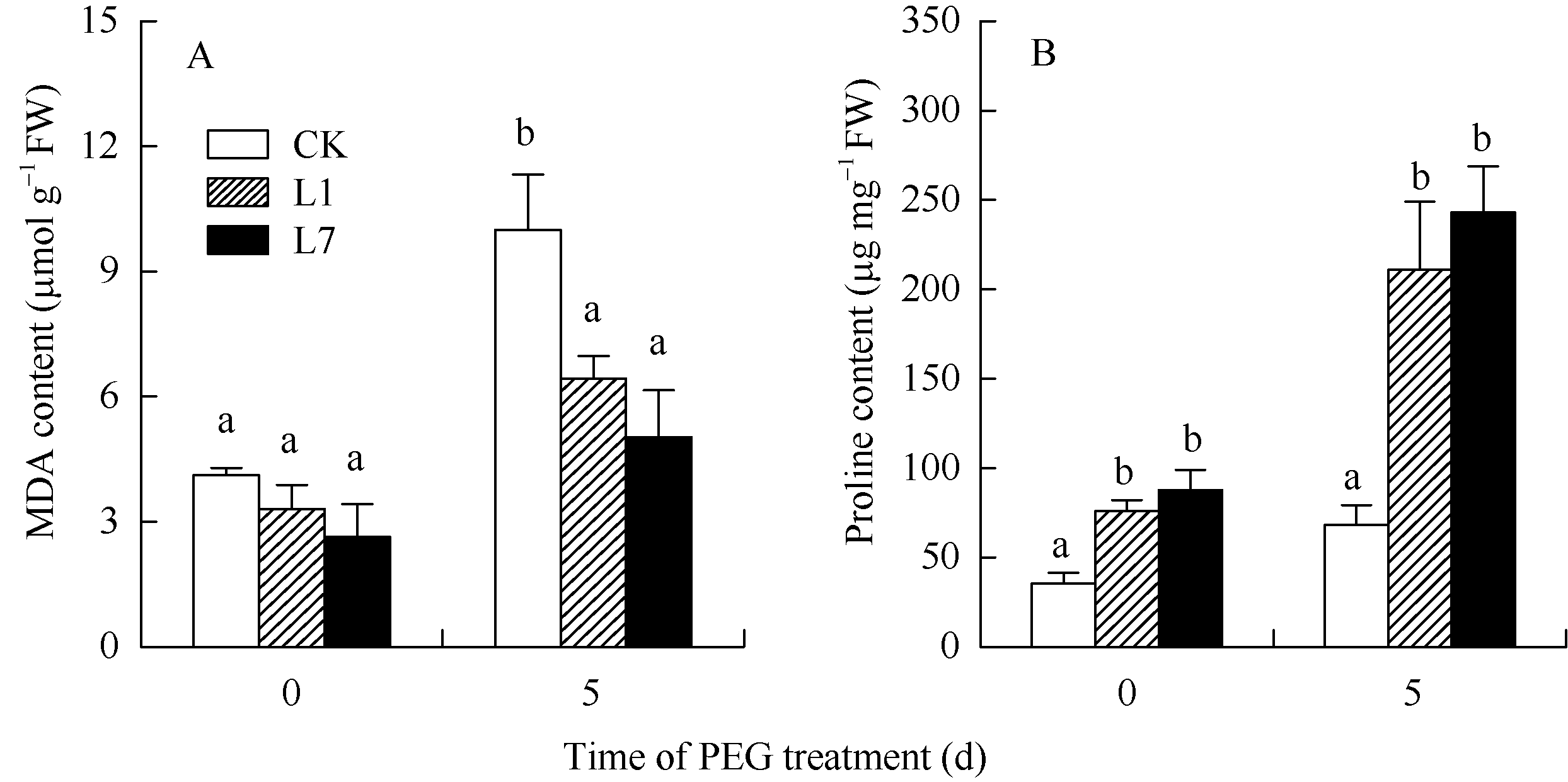
关键词:马铃薯; HD-Zip; ATHB12基因; 根; 丙二醛; 脯氨酸 Cloning and Functional Identification of the ATHB12Gene of HD-Zip IFamily in Potato ( Solanum tuberosum L.) WU Liang-Liang, YAO Lei, MA Rui, ZHU Xi, YANG Jiang-Wei, ZHANG Ning*, SI Huai-Jun Gansu Key Laboratory of Crop Genetic and Germplasm Enhancement, Gansu Provincial Key Laboratory of Aridland Crop Science / College of Life Science and Technology, Gansu Agricultural University, Lanzhou 730070, China Fund:This study was supported by the National Natural Science Foundation of China (31460370), Specialized Research Fund for the Doctoral Program of Higher Education of China (20126202110007), International Science & Technology Cooperation Program of China (0102014DFG31570), and Gansu Key Laboratory of Aridland Crop Science of Gansu Agricultural University (GSCS-2012-02) AbstractHD-Zip I is a class of plant-specific transcription factors, which has an important role in response to adversity stress in plant. A ATHB12 gene of HD-Zip I transcription factors was cloned from potato cultivar Gannongshu 2, which contains a 759 bp open reading frame (ORF) encoding a protein of 252 amino acid residues. ATHB12 gene is located on potato chromosome 1, and its promoter region sequence contains cis-acting elements including ABRE, LTRECOREATCOR15, WBOXATNPR1 responsive to abiotic stresses (ABA, temperature, dehydration, and salt stress). ATHB12 gene expressed in root, stem and leaf of potato, with the highest expression in the root. qRT-PCR analysis confirmed that the gene was induced by PEG, NaCl, and ABA, but repressed by cold treatment. The overexpressed-vector of ATHB12 gene driven by the constitutive promoter CaMV 35S was constructed, and the transgenic plants were obtained using Agrobacterium-mediated transformation system. The malondialdehyde (MDA) content in the transgenic plant leaves was significantly lower ( P< 0.05), whereas the proline content was significantly higher ( P<0.05) than those of non-transgenic control under drought stress. The fresh and dry weight of the transgenic plant root was higher than that of non-transgenic plants. These results showed that ATHB12 gene may be involved in response to stress.
图2 马铃薯ATHB12氨基酸序列与其他物种同源氨基酸序列多重比对结果 下画线: HD结构域; 虚线: LZ基序。Fig. 2 Alignment of the potato ATHB12 amino acid sequence and its homologous amino acid sequences from other species Underline: HD domain; Dot line: LZ motif.
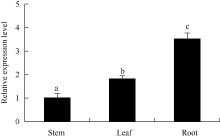
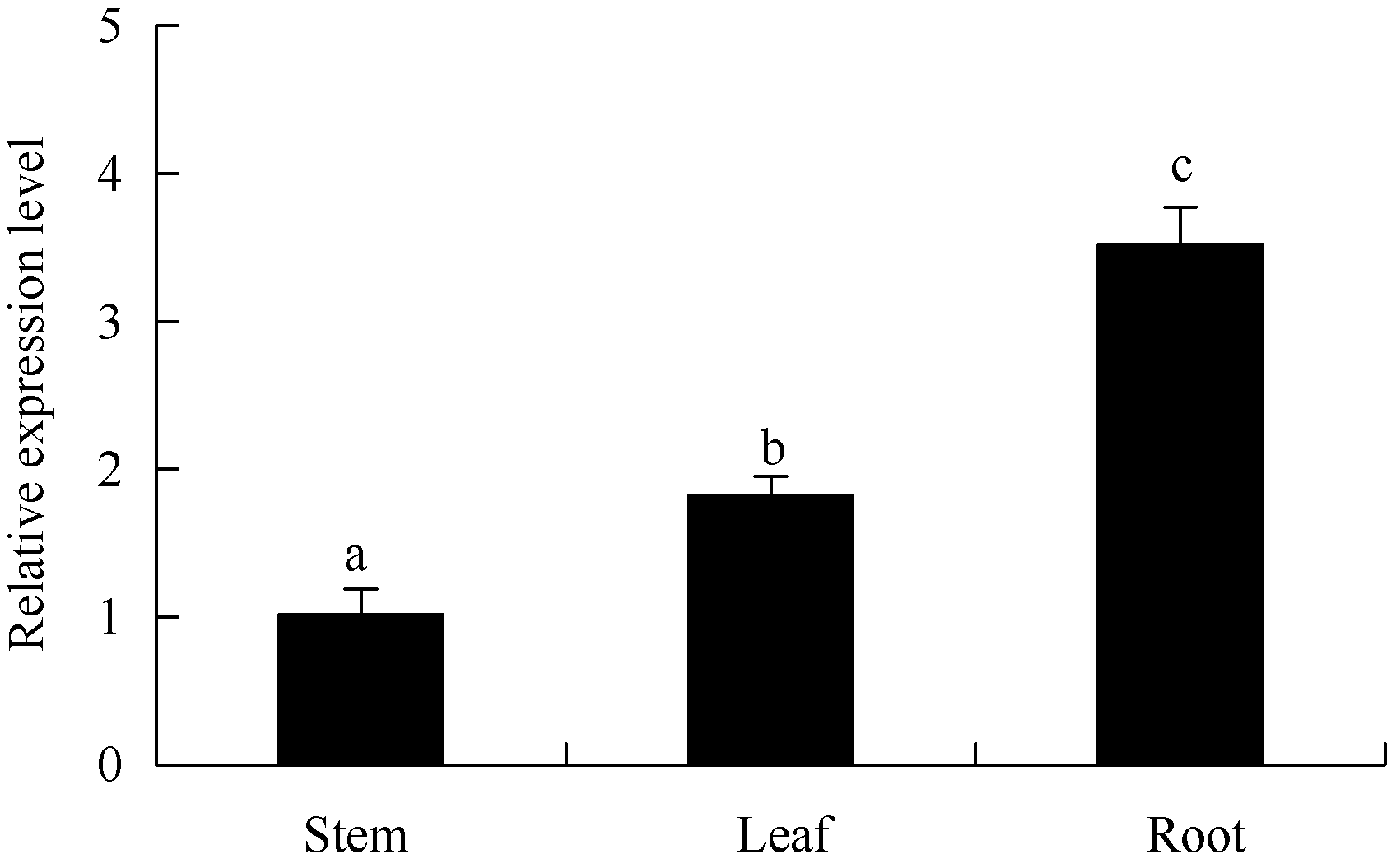
图3 马铃薯ATHB12基因在不同器官中的qRT-PCR表达检测内参基因: ef1a, n=3; 误差线表示3个重复间的标准偏差; 柱上不同字母表示差异显著(P< 0.05), 图4同。Fig. 3 qRT-PCR assay ofATHB12 gene in different organs of potatoReference gene: ef1a, n=3; Error bars indicate standard deviation for three replicates; Bars superscripted by different letters are significantly different at P< 0.05. The same as in Fig. 4.
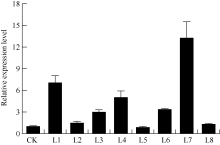
图8 转基因植株的丙二醛和脯氨酸含量 A: 丙二醛含量, B: 脯氨酸含量; CK: 非转基因植株; L1, L7: 转基因植株。数据为5次生物学重复的平均值± 标准偏差, 柱上不同字母表示差异显著(P< 0.05)。Fig. 8 MDA and proline content in the transgenic potato plants A: MDA content, B: proline content; CK: non-transgenic plants; L1, L7: transgenic plants. Values are the mean ± SD of five biological replicates. Bars superscripted by different letters are significantly different at P< 0.05.
表2 Table 2 表2(Table 2)
表2 转基因植株的丙二醛和脯氨酸含量相对值 Table 2 Relative value of MDA and proline content in the transgenic potato plants
相对值(PEG处理后/未处理) Relative value (PEG treated/untreated)
CK
L1
L7
丙二醛MDA
2.43± 0.31 a
1.99± 0.34 a
1.95± 0.23 a
脯氨酸Proline
1.95± 0.26 a
2.76± 0.22 b
2.81± 0.40 b
Values followed by different letter within the same line are significant by different at P< 0.05; n = 5. 同行中具有不同字母的平均值间差异显著(P< 0.05); n = 5。
表2 转基因植株的丙二醛和脯氨酸含量相对值 Table 2 Relative value of MDA and proline content in the transgenic potato plants
表3 转基因植株试管苗株高和根重 Table 3 Plant height and root weight of the transgenic test-tube plant
株系Line
株高Plant height (cm)
根鲜重Root fresh weight (mg)
根干重Root dry weight (mg)
CK
6.0± 0.77 a
20.9± 1.5 a
1.3± 0.38 a
L1
6.2± 1.20 a
35.4± 2.3 b
2.7± 0.74 b
L7
6.5± 0.93 a
37.8± 4.2 c
3.2± 0.83 b
Values followed by different letter within the same line are significant different at P< 0.05. Statistics related indicators were calculated after three weeks of plantlets subculture. The data represent the mean ± standard deviation of five biological replicates. 同列中具有不同字母的平均值间差异显著(P< 0.05); 试管苗转接3周后统计相关指标。数据为5次生物学重复的平均值± 标准偏差。
表3 转基因植株试管苗株高和根重 Table 3 Plant height and root weight of the transgenic test-tube plant
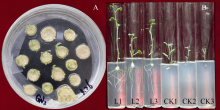
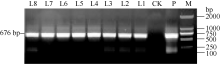
图9 转基因植株试管苗表型差异 CK: 非转基因植株; L1, L7: 转基因植株。Fig. 9 Phenotype difference between the transgenic test-tube plants and the control CK: non-transgenic plants; L1, L7: transgenic plants
Ariel FD, Manavella PA, Dezar CA, Chan RL. The true story of the HD-Zip family. Trends Plant Sci, 2007, 12: 419-426[本文引用:4]
[2]
De SmetI, LauS, Ehrismann JS, AxiotisI, KolbM, KientzM, WeijersD, JurgensG. Transcriptional repression of BODENLOS by HD-ZIP transcription factor HB5 in Arabidopsis thaliana. J Exp Bot, 2013, 64: 3009-3019[本文引用:1]
[3]
Re DA, Dezar CA, Chan RL, Baldwin IT, BonaventureG. Nicotiana attenuata NaHD20 plays a role in leaf ABA accumulation during water stress, benzylacetone emission from flowers, and the timing of bolting and flower transitions. J Exp Bot, 2011, 62 : 155-166[本文引用:1]
[4]
ArielF, DietA, VerdenaudM, GruberV, FrugierF, ChanR, CrespiM. Environmental regulation of lateral root emergence in Medicago truncatula requires the HD-Zip I transcription factor HB1. Plant Cell, 2010, 22: 2171-2183[本文引用:2]
[5]
SakakibaraK, NishiyamaT, KatoM, HasebeM. Isolation of homeodomain-leucine zipper genes from the moss Physcomitrella patens and the evolution of homeodomain-leucine zipper genes in land plants. Mol Biol Evol, 2001, 18: 491-502[本文引用:1]
[6]
SodermanE, HjellstromM, FahlesonJ, EngstromP. The HD-Zip gene ATHB6 in Arabidopsis is expressed in developing leaves, roots and carpels and up-regulated by water deficit conditions. Plant Mol Biol, 1999, 40: 1073-1083[本文引用:1]
[7]
Olsson AS, EngstromP, SodermanE. The homeobox genes ATHB12 and ATHB7 encode potential regulators of growth in response to water deficit in Arabidopsis. Plant Mol Biol, 2004, 55: 663-677[本文引用:3]
[8]
DengX, PhillipsJ, Meijer AH, SalaminiF, BartelsD. Characterization of five novel dehydration-responsive homeodomain leucine zipper genes from the resurrection plant Craterostigma plantagineum. Plant Mol Biol, 2002, 49: 601-610[本文引用:1]
[9]
Dezar CA, Gago GM, Gonzalez DH, Chan RL. Hahb-4, a sunflower homeobox-leucine zipper gene, is a developmental regulator and confers drought tolerance to Arabidopsis thaliana plants. Transgenic Res, 2005, 14: 429-440[本文引用:1]
[10]
ZhaoY, MaQ, JinX, PengX, LiuJ, DengL, YanH, ShengL, JiangH, ChengB. A novel maize homeodomain-leucine zipper (HD-Zip) I gene, Zmhdz10positively regulates drought and salt tolerance in both rice and Arabidopsis. Plant Cell Physiol, 2014, 55: 1142-1156[本文引用:2]
[11]
Potato Genome Sequencing Consortium, Xu X, Pan S, Cheng S, Zhang B, Mu D, Ni P, Zhang G, Yang S, Li R, Wang J, Orjeda G, Guzman F, Torres M, Lozano R, Ponce O, Martinez D, De la Cruz G, Chakrabarti S K, Patil V U, Skryabin K G, Kuznetsov B B, Ravin N V, Kolganova T V, Beletsky A V, Mardanov A V, Di Genova A, Bolser D M, Martin D M, Li G, Yang Y, Kuang H, Hu Q, Xiong X, Bishop G J, Sagredo B, Mejía N, Zagorski W, Gromadka R, Gawor J, Szczesny P, Huang S, Zhang Z, Liang C, He J, Li Y, He Y, Xu J, Zhang Y, Xie B, Du Y, Qu D, Bonierbale M, Ghislain M, Herrera Mdel R, Giuliano G, Pietrella M, Perrotta G, Facella P, O'Brien K, Feingold S E, Barreiro L E, Massa G A, Diambra L, Whitty B R, Vaillancourt B, Lin H, Massa A N, Geoffroy M, Lundback S, DellaPenna D, Buell C R, Sharma S K, Marshall D F, Waugh R, Bryan G J, Destefanis M, Nagy I, Milbourne D, Thomson S J, Fiers M, Jacobs J M, Nielsen K L, Sønderkær M, Iovene M, Torres G A, Jiang J, Veilleux R E, Bachem C W, de Boer J, Borm T, Kloosterman B, van Eck H, Datema E, Hekkert B L, Goverse A, van Ham R C, Visser R G. Genome sequence and analysis of the tuber crop potato. Nature, 2011, 475: 189-195[本文引用:1]
[12]
张宁, 司怀军, 王蒂. 拟南芥rd29A基因启动子克隆及其在马铃薯抗胁迫转基因中的应用. 作物学报, 2005, 31: 159-164ZhangN, Si HJ, WangD. Cloning of rd29A gene promoter from Arabidopsis thaliana and its application in stress-resistance transgenic potato. Acta Agron Sin, 2005, 31: 159-164 (in Chinese with English abstract)[本文引用:2]
[13]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2 (-Delta Delta C(T)) method. Methods, 2001, 25: 402-408[本文引用:1]
[14]
司怀军, 谢从华, 柳俊. 农杆菌介导的马铃薯试管薯遗传转化体系的优化及反义Class Ipatatin基因的导入. 作物学报, 2003, 29: 801-805Si HJ, Xie CH, LiuJ. An efficient protocol for Agrobacterium-mediated transformation with microtuber and the introduction of an antisense class I patatin gene into potato. Acta Agron Sin, 2003, 29: 801-805 (in Chinese with English abstract)[本文引用:1]
[15]
EdwardsK, JohnstoneC, ThompsonC. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucl Acids Res, 1991, 19: 1349-1352[本文引用:1]
[16]
陈建勋, 王晓峰. 植物生理学实验指导. 广州: 华南理工大学出版社, 2002. pp 119-124Chen JX, Wang XF. Experiment Manual for Plant Physiology. Guangzhou: South China University of Technology Press, 2002. pp 119-124(in chinese)[本文引用:1]
[17]
Prigge MJ, OtsugaD, Alonso JM, Ecker JR, Drews GN, Clark SE. Class III homeodomain-leucine zipper gene family members have overlapping, antagonistic, and distinct roles in Arabidopsis development. Plant Cell, 2005, 17: 61-76[本文引用:1]
[18]
WangY, HenrikssonE, SodermanE, Henriksson KN, SundbergE, EngstromP. The Arabidopsis homeobox gene, ATHB16, regulates leaf development and the sensitivity to photoperiod in Arabidopsis. Dev Biol, 2003, 264: 228-239[本文引用:1]
[19]
CapellaM, Ribone PA, Arce AL, Chan RL. Arabidopsis thaliana HomeoBox 1 (AtHB1), a Homedomain-Leucine Zipper I (HD-Zip I) transcription factor, is regulated by PHYTOCHROME- INTERACTING FACTOR 1 to promote hypocotyl elongation. New Phytol, 2015, 207: 669-682[本文引用:2]
[20]
ZhangS, HaiderI, KohlenW, JiangL, BouwmeesterH, Meijer AH, SchluepmannH, Liu CM, Ouwerkerk PB. Function of the HD-Zip I gene Oshox22 in ABA-mediated drought and salt tolerances in rice. Plant Mol Biol, 2012, 80: 571-585[本文引用:1]
[21]
HuT, YeJ, TaoP, LiH, ZhangJ, ZhangY, YeZ. Tomato HD-Zip I transcription factor, SlHZ24, modulates ascorbate accumulation through positively regulating the D-mannose/L-galactose pathway. Plant J, 2016, 85: 16-29[本文引用:1]
[22]
LiuW, FuR, LiQ, LiJ, WangL, RenZ. Genome-wide identification and expression profile of homeodomain-leucine zipper Class I gene family in Cucumis sativus. Gene, 2013, 531: 279-287[本文引用:2]
[23]
梁毅, 刘小义, 张洪伟, 谭武平. 洋葱花青素合成相关基因(AcPAL1)的克隆和表达分析. 农业生物技术学报, 2014, 22: 47-54LiangY, Liu XY, Zhang HW, Tan WP. Cloning and expression analysis of an anthocyanin bio-synthesis-related gene (AcPAL1) in onion (Allium cepa L. ). J Agric Biotechnol, 2014, 22: 47-54 (in Chinese with English abstract)[本文引用:1]
[24]
亢键, 姜永华, 王豪杰, 杨艳青, 任小林. 苹果果实HD-Zip I转录因子亚家族基因鉴定及表达分析. 西北农业学报, 2014, 23: 160-165KangJ, Jiang YH, Wang HJ, Yang YQ, Ren XL. Identification and expression analysis of HD-ZIP Itranscription factor genes in apple fruit. Acta Bot Boreali-Occident Sin, 2014, 23: 160-165 (in Chinese with English abstract)[本文引用:1]
[25]
TaoZ, KouY, LiuH, LiX, XiaoJ, WangS. OsWRKY45 alleles play different roles in abscisic acid signalling and salt stress tolerance but similar roles in drought and cold tolerance in rice. J Exp Bot, 2011, 62: 4863-4874[本文引用:1]
[26]
SongY, WangL, XiongL. Comprehensive expression profiling analysis of OsIAA gene family in developmental processes and in response to phytohormone and stress treatments. Planta, 2009, 229: 577-591[本文引用:1]
[27]
LuanM, XuM, LuY, ZhangL, FanY, WangL. Expression of zma-miR169 miRNAs and their target ZmNF-YA genes in response to abiotic stress in maize leaves. Gene, 2015, 555: 178-185[本文引用:1]
[28]
Hou XJ, Li SB, Liu SR, Hu CG, Zhang JZ. Genome-wide classification and evolutionary and expression analyses of citrus MYB transcription factor families in sweet orange. PloS One, 2014, 9: e95489[本文引用:1]
[29]
DaiM, HuY, MaQ, ZhaoY, Zhou DX. Functional analysis of rice HOMEOBOX4 (Oshox4) gene reveals a negative function in gibberellin responses. Plant Mol Biol, 2008, 66: 289-301[本文引用:1]
[30]
惠非琼, 彭兵, 楼兵干, 林福呈, 聂长春, 刘剑. 印度梨形孢通过促进渗透调节物质的合成和诱导抗逆相关基因的表达提高烟草耐盐性. 农业生物技术学报, 2014, 22: 168-176Hui FQ, PengB, Lou BG, Lin FC, Nie CC, LiuJ. Piriformospora indica improves salt tolerance in Nicotiana tobacum by promoting the synthesis of osmolyte and inducing the expression of stress resistance genes. J Agric Biotechnol, 2014, 22: 168-176 (in Chinese with English abstract)[本文引用:2]
 , 姚磊, 马瑞, 朱熙, 杨江伟, 张宁
, 姚磊, 马瑞, 朱熙, 杨江伟, 张宁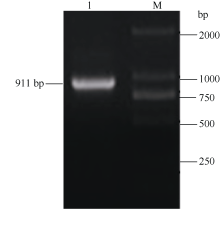








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}