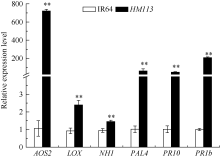
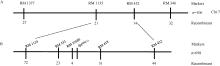
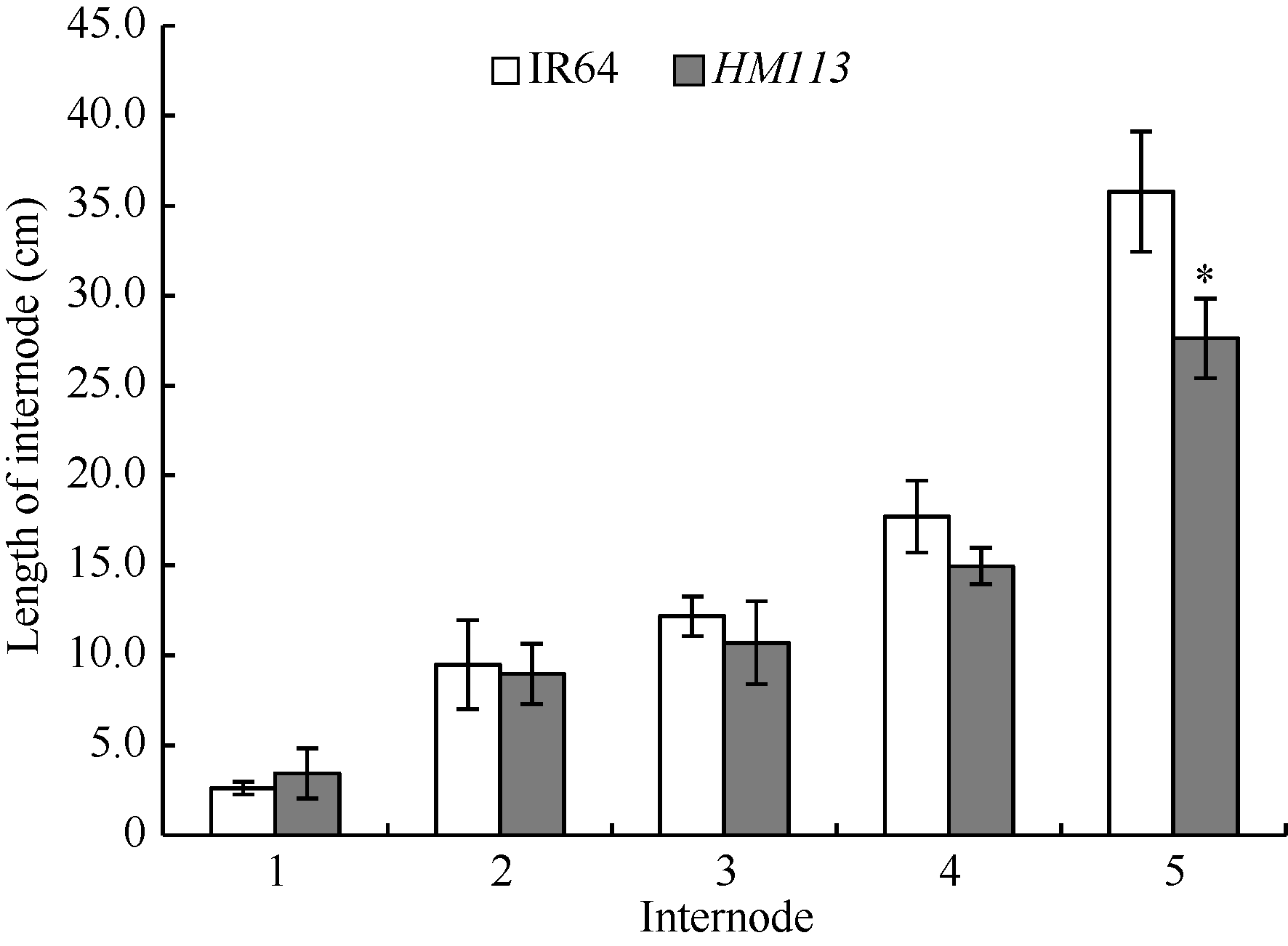
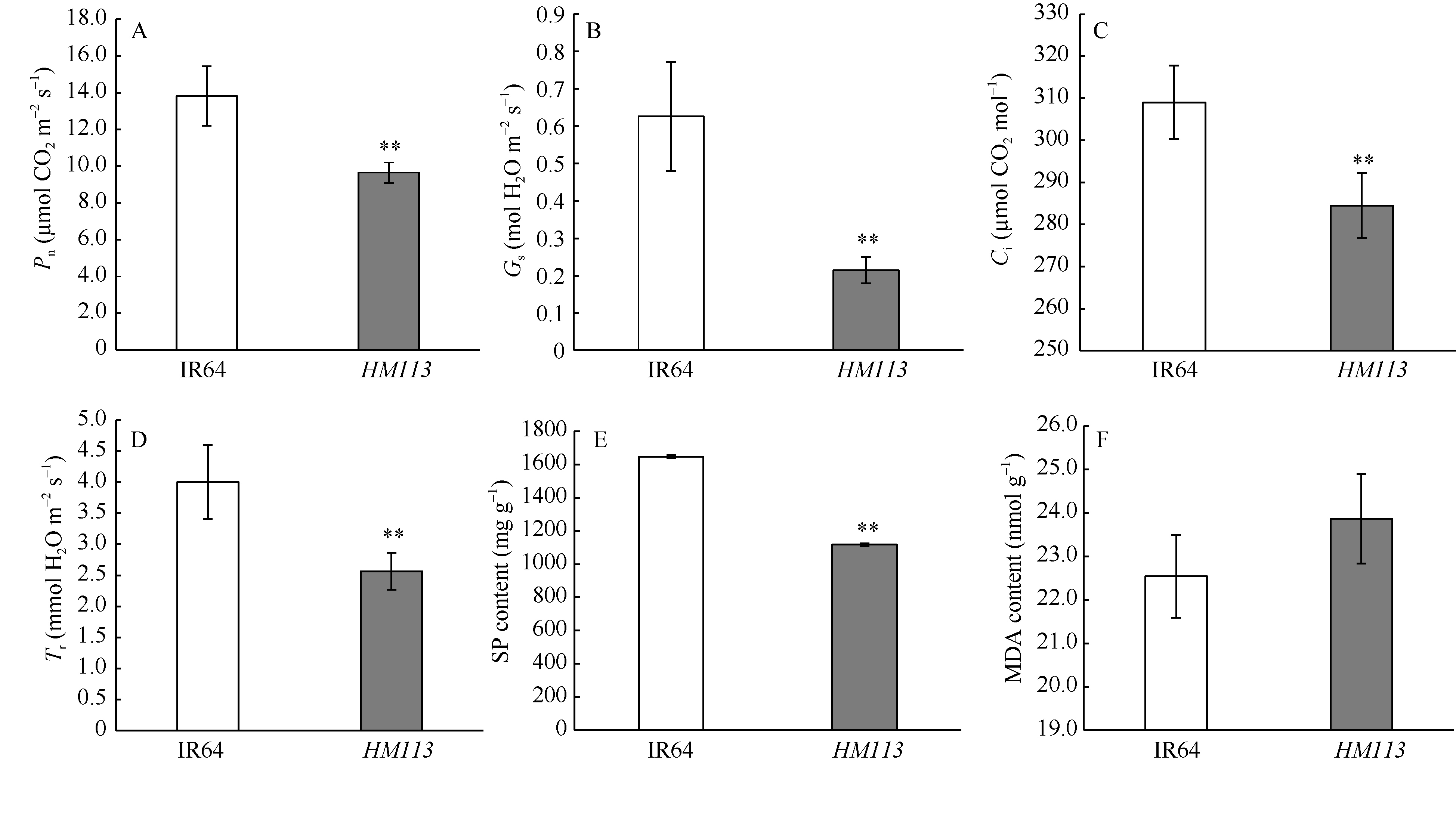
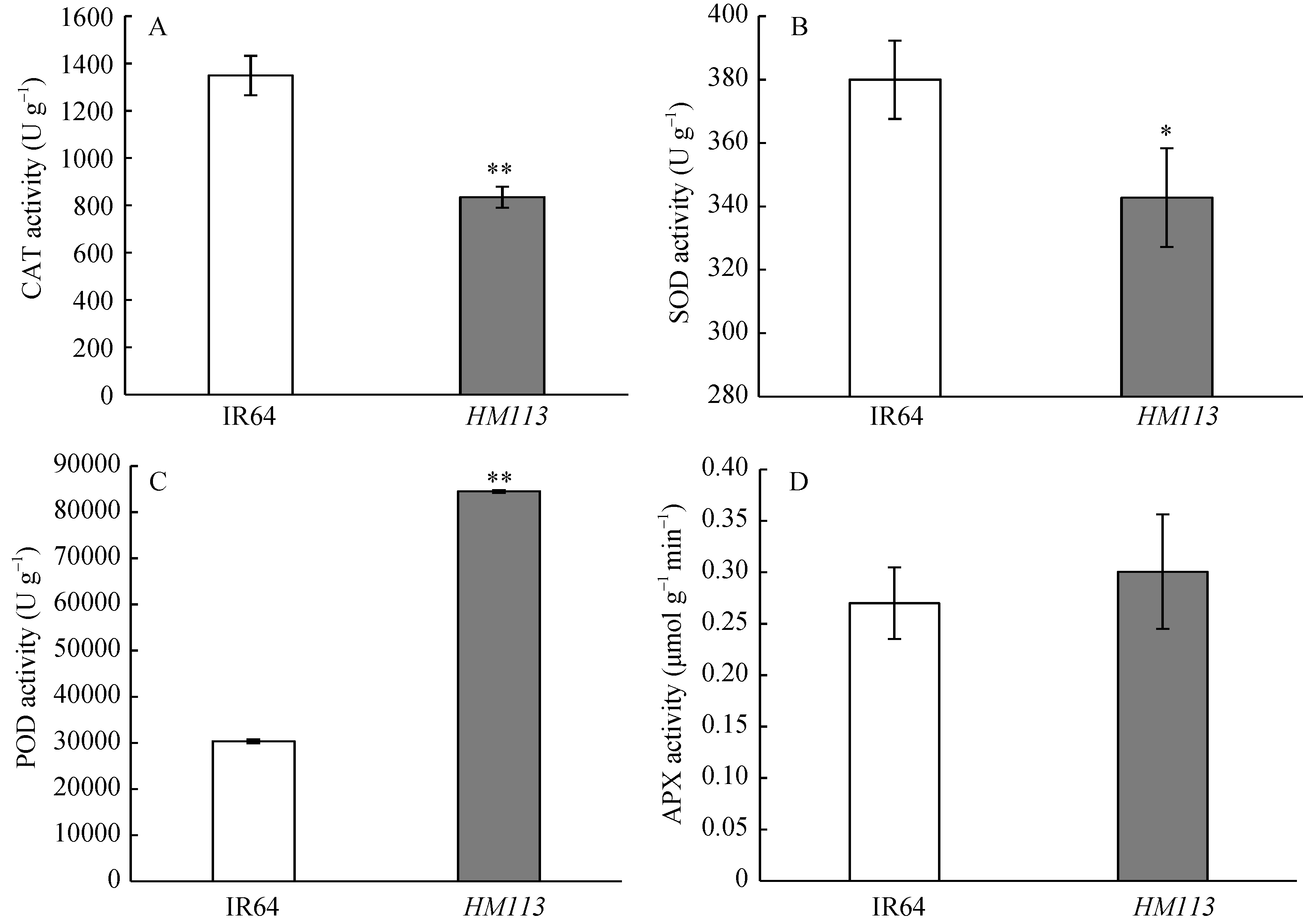
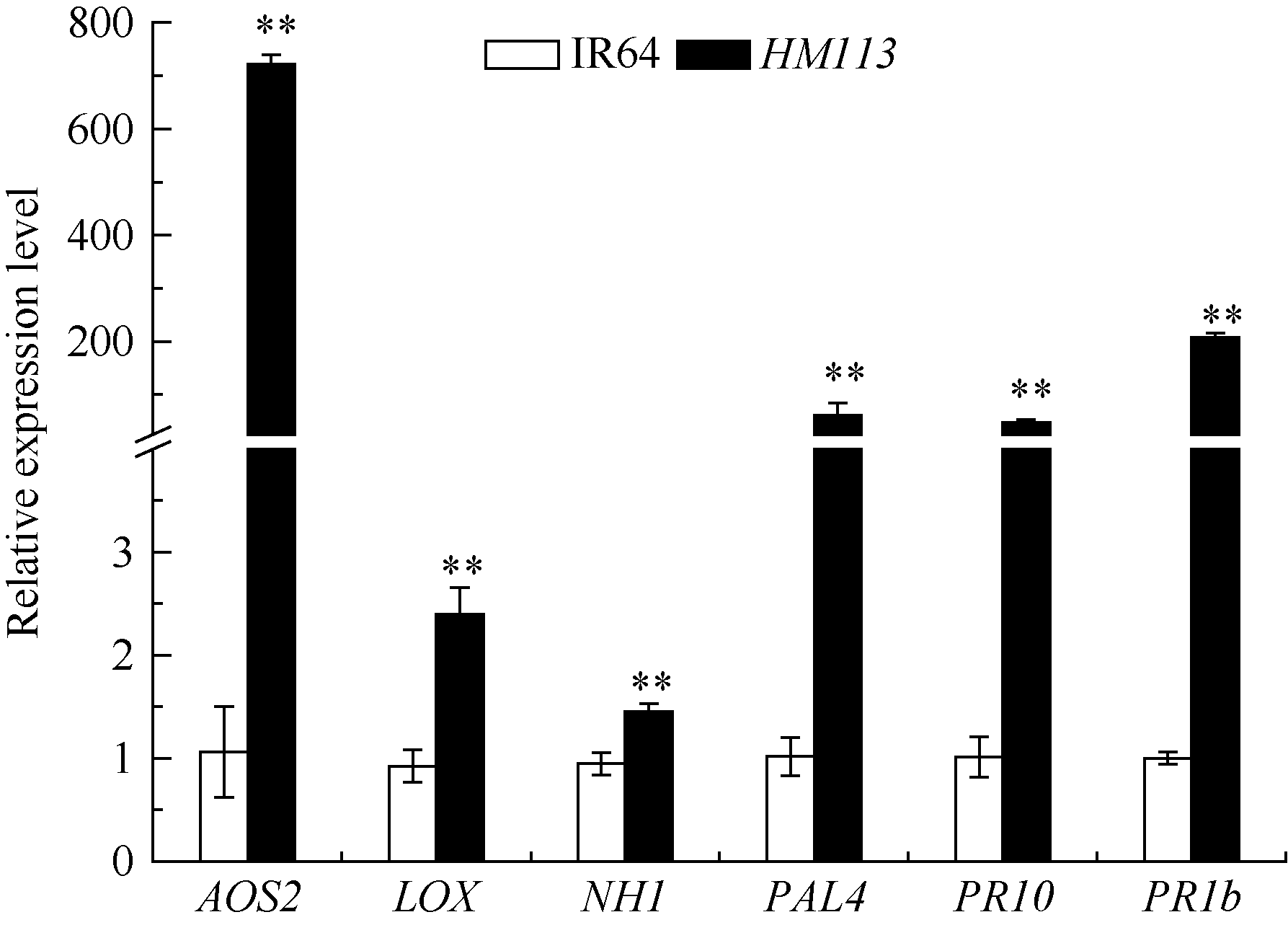
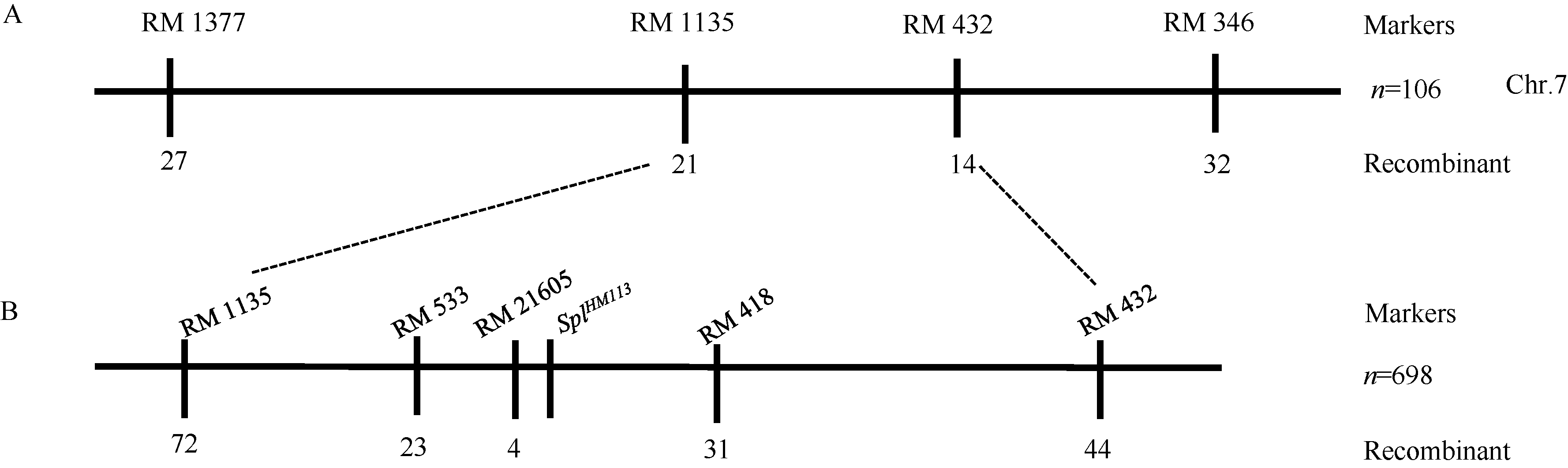
关键词:水稻; 斑点叶突变体; 白叶枯病抗性; 活性氧; 基因定位 Characterization and Gene Fine Mapping of a Rice Dominant Spotted-leaf Mutant GUO Dan, SHI Yong-Feng, WANG Hui-Mei, ZHANG Xiao-Bo, SONG Li-Xin, XU Xia, HE Yan, GUO Liang, WU Jian-Li* State Key Laboratory of Rice Biology / Chinese National Center for Rice Improvement / China National Rice Research Institute, Hangzhou 310006, China Fund:This study was supported by the Natural Science Foundation of Zhejiang Province (LQ15C130005) and the National High-tech R&D Program of China (2014AA10A603-15) AbstractA stable inherited rice spotted-leaf mutant HM113 was isolated from an EMS-induced IR64 mutant bank. Under natural conditions, brown lesions were observed on the leaves in three weeks after sowing and spread to the sheaths at the initial heading stage. Agronomic traits including the plant height, panicle length, number of panicles, number of filled grain/panicle, seed-setting rate and 1000-grain weight were decreased significantly in HM113. In addition, the photosynthetic pigment contents, net photosynthetic rate and soluble protein content in the mutant were significantly lower than those in the wild type IR64, while the MDA content was similar to that in the wild-type. Activities of CAT and SOD were significantly lower and activity of POD was significantly higher in the mutant than in IR64. Histochemical analysis showed that cell death and ROS accumulation were occurred in and around the lesions in HM113. Furthermore, disease resistance to bacterial blight pathogens was significantly enhanced in the mutant in contrast to that in the wild type IR64. Expression of defense-related genes including AOS2, PAL4, PR10, and PR1b was apparently up-regulated in the mutant. Genetic analysis indicated that the mutant trait was controlled by a novel single dominant nuclear gene, tentatively termed as SplHM113, which was detected to be located in a region around 308 kb flanked by RM21605 and RM418 on the long arm of chromosome 7. The data and populations obtained in the present study would facilitate the isolation and functional analysis of SplHM113.
Keyword:Rice; Spotted-leaf mutant; Bacterial blight resistance; Reactive oxygen species; Gene mapping Show Figures Show Figures
图5 突变体HM113的组织化学染色 A: IR64苔盼蓝染色前; B: IR64苔盼蓝染色后; C: HM113苔盼蓝染色前; D: HM113苔盼蓝染色后; E: IR64 DAB染色前; F: IR64 DAB染色后; G: HM113 DAB染色前; H: HM113 DAB染色后; I: IR64 NBT染色前; J: IR64 NBT染色后; K: HM113NBT染色前; L: HM113NBT染色后。Fig. 5 Histochemical analysis of the mutant HM113 A: IR64 before trypan blue staining; B: IR64 after trypan blue staining; C:HM113 before trypan blue staining; D:HM113 after trypan blue staining; E: IR64 before DAB staining; F: IR64 after DAB staining; G: HM113 before DAB staining; H: HM113after DAB staining; I: IR64 before NBT staining; J: IR64 after NBT staining; K: HM113 before NBT staining; L: HM113 after NBT staining.
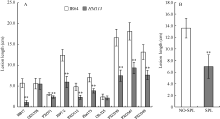
2.6 突变体HM113对白叶枯病菌的抗性增强 与野生型相比, 突变体对菌株HB17、PXO71、JS97-2、PXO112、Zhe173、PXO339、PXO347和PXO349抗性极显著增强; 对菌株GD1358和OS-225的抗性则无明显差异(图7-A)。选用菌株PXO349接种HM113/IR64回交F2群体, 其中184个有斑植株的病斑平均长度为6.96 cm ± 2.04 cm, 极显著低于65个无斑植株的13.58 cm ± 1.07 cm (图7-B)。说明突变体对白叶枯病菌的抗性普遍增强, 且抗性增强与褐斑的存在高度相关。 图6 Fig. 6
图7 突变体HM113的白叶枯病抗性 A: IR64和HM113的白叶枯病抗性反应; B: HM113/IR64的F2群体对PXO349的抗性反应。NO-SPL: 无斑单株; SPL: 有斑单株。* * 极显著差异(P≤ 0.01)。Fig. 7 Resistance of mutant HM113 to Xanthomonas oryzae pv. oryzae(Xoo) A: reaction of IR64 and HM113 to Xoo; B: reaction of F2 individuals derived from IR64/HM113 to PXO349. NO-SPL: no-spotted plant; SPL: spotted-plant. * * Significantly different at P ≤ 0.01.
HuG, YalpaniN, Briggs SP, Johal GS. A porphyrin pathway impairment is responsible for the phenotype of a dominant disease lesion mimic mutant of maize. Plant Cell, 1998, 10: 1095-1105[本文引用:1]
[5]
BalagueC, LinB, AlconC, FlottesG, MalmstromS, KohlerC, NeuhausG, PelletierG, GaymardF, RobyD. HLM1, an essential signaling component in the hypersensitive response, is a member of the cyclic nucleotide-gated channel ion channel family. Plant Cell, 2003, 15: 365-379[本文引用:1]
[6]
Feng BH, YangY, Shi YF, Shen HC, Wang HM, Huang QN, XuX, Lv XG, Wu JL. Characterization and genetic analysis of a novel rice spotted-leaf mutant HM47 with broad-spectrum resistance to Xanthomonas oryzae pv. oryzae. J Integr Plant Biol, 2013, 55: 473-483[本文引用:2]
[7]
Shen HC, Shi YF, Feng BH, Wang HM, XuX, Huang QN, Lv XG, Wu JL. Identification and genetic analysis of a novel rice spotted-leaf mutant with broad-spectrum resistance to Xanthomonas oryzae pv. oryzae. J Integr Agric, 2014, 13: 713-721[本文引用:1]
[8]
李小红, 施勇烽, 张晓波, 奉保华, 宋莉欣, 王惠梅, 徐霞, 黄奇娜, 郭丹, 吴建利. 水稻斑点叶突变体hm197的鉴定及其基因定位. 中国水稻科学, 2015, 29: 447-456Li XH, Shi YF, Zhang XB, Feng BH, Song LX, Wang HM, XuX, Huang QN, GuoD, Wu JL. Identification and gene mapping of a spotted leaf mutant hm197 in rice. Chin J Rice Sci, 2015, 29: 447-456 (in Chinese with English abstract)[本文引用:1]
[9]
Jung YH, Lee JH, Agrawal GK, RakwalR, Kim JA, Shim JK, Lee SK, Jeon JS, Koh HJ, Lee YH, IwahashiH, Jwa NS. The rice (Oryza sativa) blast lesion mimic mutant, blm, may confer resistance to blast pathogens by triggering multiple defense- associated signaling pathways. Plant Physiol Biochem, 2005, 43: 397-406[本文引用:1]
[10]
WuC, BordeosA, Madamba MR, BaraoidanM, RamosM, Wang GL, Leach JE, LeungH. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics, 2008, 279: 605-619[本文引用:1]
[11]
FujiwaraT, MaisonneuveS, IsshikiM, MizutaniM, ChenL, Wong HL, KawasakiT, ShimamotoK. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J Biol Chem, 2010, 285: 11308-11313[本文引用:1]
[12]
WangL, PeiZ, TianY, HeC. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol Plant Microbe Interact, 2005, 18: 375-384[本文引用:1]
[13]
SunC, LiuL, TangJ, LinA, ZhangF, FangJ, ZhangG, ChuC. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J Genet Genomics, 2011, 38: 29-37[本文引用:1]
[14]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1]
[15]
YamanouchiU, YanoM, LinH, AshikariM, YamadaK. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA, 2002, 99: 7530-7535[本文引用:1]
[16]
Jiang CJ, ShimonoM, MaedaS, InoueH, MoriM, HasegawaM, SuganoS, TakatsujiH. Suppression of the rice fatty-acid desaturase gene OsSSI2 enhances resistance to blast and leaf blight diseases in rice. Mol Plant Microbe Interact, 2009, 22: 820-829[本文引用:1]
[17]
QiaoY, JiangW, LeeJ, ParkB, Choi MS, PiaoR, Woo MO, Roh JH, HanL, Paek NC, Seo HS, Koh HJ. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:3]
[18]
FekihR, TamiruM, KanzakiH, AbeA, YoshidaK, KanzakiE, SaitohH, TakagiH, NatsumeS, Undan JR, UndanJ, TerauchiR. The rice (Oryza sativa L. ) LESION MIMIC RESEMBLING, which encodes an AAA-type ATPase, is implicated in defense response. Mol Genet Genomics, 2015, 290: 611-622[本文引用:1]
[19]
Arnon DI. Copper enzymes in isolated chloroplasts polyphenoloxidase in Beta vulgaris. Plant Physiol, 1949, 24: 1-15[本文引用:1]
[20]
Wellburn AR. The spectral determination of chlorophyll a and b, as well as total carotenoids, using various solvents with spectro-photometers of different resolution. Plant Physiol, 2015, 144: 307-313[本文引用:1]
[21]
赵世杰, 史国安, 董新纯. 植物生理学实验指导. 北京: 中国农业科学技术出版社, 2002. pp 134-143Zhao SJ, Shi GA, Dong XC. Plant Physiology Experiment Instruction. Beijing: China Agricultural Science and Technology Press, 2002. pp 134-143(in Chinese)[本文引用:2]
[22]
Huang QN, Shi YF, Zhang XB, Song LX, Feng BH, Wang HM, XuX, Li XH, GuoD, Wu JL. Single base substitution in OsCDC48 is responsible for premature senescence and death phenotype in rice. J Integr Plant Biol, 2016, 58: 12-28[本文引用:1]
[23]
YinZ, ChenJ, ZengL, GohM, LeungH, Khush GS, Wang GL. Characterizing rice lesion mimic mutants and identifying a mutant with broad-spectrum resistance to rice blast and bacterial blight. Mol Plant Microbe Interact, 2000, 13: 869-876[本文引用:1]
[24]
Kauffman HE, Reddy A P D, Ksiek S P V, Marca S D. An improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis Rep, 1973, 57: 537-541[本文引用:1]
[25]
卢扬江, 郑康乐. 提取水稻DNA的一种简易方法. 中国水稻科学, 1992, 6: 47-48Lu YJ, Zheng KL. A simple method for isolation of rice DNA. Chin J Rice Sci, 1992, 6: 47-48 (in Chinese with English abstract)[本文引用:1]
[26]
Shi YF, ChenJ, Liu WQ, Huang QN, ShenB, LeungH, Wu JL. Genetic analysis and gene mapping of a new rolled-leaf mutant in rice (Oryza sativa L. ). Sci China Ser C: Life Sci, 2009, 52: 885-890[本文引用:1]
[27]
代高猛, 朱小燕, 李云峰, 凌英华, 赵芳明, 杨正林, 何光华. 水稻类病斑突变体spl31的遗传分析与基因定位. 作物学报, 2013, 39: 1223-1230Dai GM, Zhu XY, Li YF, Ling YH, Zhao FM, Yang ZL, He GH. Genetic analysis and fine mapping of a lesion mimic mutant spl31 in rice. Acta Agron Sin, 2013, 39: 1223-1230 (in Chinese with English abstract)[本文引用:1]
[28]
宋莉欣, 黄奇娜, 奉保华, 施勇烽, 张晓波, 徐霞, 王惠梅, 李小红, 赵宝华, 吴建利. 水稻斑点叶突变体spl21的鉴定与基因定位. 作物学报, 2015, 41: 1519-1528Song LX, Huang QN, Feng BH, Shi YF, Zhang XB, XuX, Wang HM, Li XH, Zhao BH, Wu JL. Characterization and gene mapping of a spotted-leaf mutant spl21 in rice (Oryza sativa L. ). Acta Agron Sin, 2015, 41: 1519-1528 (in Chinese with English abstract)[本文引用:1]
[29]
Xiao GQ, Zhang HW, Lu XY, Huang RF. Characterization and mapping of a novel light-dependent lesion mimic mutant lmm6 in rice (Oryza sativa L. ). J Integr Agric, 2015, 14: 1687-1696[本文引用:1]
[30]
Jacks ThomasJ, Davidonis GayleH. Superoxide, hydrogen peroxide, and the respiratory burst of fungally infected plant cells. Mol Cellular Biochem, 1996, 158: 77-79[本文引用:1]
MinibayevaF, Beckett RP, KrannerI. Roles of apoplastic peroxidases in plant response to wounding. Phytochemistry, 2015, 112: 122-129[本文引用:1]
[33]
KawanoT. Roles of the reactive oxygen species-generating peroxidase reactions in plant defense and growth induction. Plant Cell Rep, 2003, 21: 829-837[本文引用:1]
[34]
WhitakerC, Beckett RP, Minibayevab FV, KrannerI. Production of reactive oxygen species in excised, desiccated and cryopreserved explants of Trichilia dregeana Sond. S Afr J Bot, 2010, 76: 112-118[本文引用:1]
[35]
金杨. 水稻类病变突变体spl5细胞坏死机制及其抗病性的研究. 浙江师范大学硕士学位论文, 浙江金华, 2009JinY. Mechanisms of Cell Death and Its Resistance in Rice Lesion Mimic Mutant spl5. MS Thesis of Zhejiang Normal University, Jinhua, China, 2009 (in Chinese with English abstract)[本文引用:1]
[36]
KaurilindE, XuE, BroscheM. A genetic framework for H2O2 induced cell death in Arabidopsis thaliana. BMC Genomics, 2015, 16: 837-853[本文引用:1]
[37]
KariolaT, BraderG, LiJ, Palva ET. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell, 2005, 17: 282-294[本文引用:1]
[38]
ChernM, Fitzgerald HA, Canlas PE, Navarre DA, Ronald PC. Overexpression of a rice NPR1 homolog leads to constitutive activation of defense response and hypersensitivity to light. Mol Plant Microbe Interact, 2005, 18: 511-520[本文引用:1]
[39]
MoriM, TomitaC, SugimotoK, HasegawaM, HayashiN, Dubouzet JG, OchiaiH, SekimotoH, HirochikaH, KikuchiS. Isolation and molecular characterization of a Spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol Biol, 2007, 63: 847-860[本文引用:2]
[40]
钟振泉, 罗文龙, 刘永柱, 王慧, 陈志强, 郭涛. 一份新的水稻斑点叶突变体spl32的鉴定和基因定位. 作物学报, 2015, 41: 861-871Zhong ZQ, Luo WL, Liu YZ, WangH, Chen ZQ, GuoT. Characterization of a novel spotted leaf mutant spl32 and mapping of Spl32(t) gene in rice (Oryza sativa). Acta Agron Sin, 2015, 41: 861-871 (in Chinese with English abstract)[本文引用:1]
[41]
Hwang SH, Hwang DJ. Isolation and characterization of the rice NPR1 promoter. Plant Biotechnol Rep, 2010, 4: 29-35[本文引用:1]
[42]
Yuan YX, Zhong SH, LiQ, Zhu ZR, Lou YG, Wang LY, Wang JJ, Wang MY, Li QL, Yang DL, He ZH. Functional analysis of rice NPR1-like genes reveals that OsNPR1/NHI is the rice orthologue conferring disease resistance with enhanced herbivore susceptibility. Plant Biotechnol J, 2007, 5: 313-324[本文引用:1]
[43]
LiZ, Zhang YX, LiuL, Liu QE, Bi ZZ, YuN, Cheng SH, Cao LY. Fine mapping of the lesion mimic and early senescence 1 (lmes1) in rice (Oryza sativa). Plant Physiol Biochem, 2014, 80: 300-307[本文引用:1]
[44]
Chen XF, Pan JW, ChengJ, Jiang GH, JinY, Gu ZM, QianQ, Zhai WX, Ma BJ. Fine genetic mapping and physical delimitation of the lesion mimic gene spotted leaf 5 (spl5) in rice (Oryza sativa L. ). Mol Breed, 2009, 24: 387-395[本文引用:1]
[45]
BabuR, Jiang CJ, XuX, Kottapalli KR, TakatsujiH, MiyaoA, HirochikaH, KawasakiS. Isolation, fine mapping and expression profiling of a lesion mimic genotype, spl(NF4050-8) that confers blast resistance in rice. Theor Appl Genet, 2011, 122: 831-854[本文引用:1]
 , 施勇烽, 王惠梅, 张晓波, 宋莉欣, 徐霞, 贺彦, 郭梁, 吴建利
, 施勇烽, 王惠梅, 张晓波, 宋莉欣, 徐霞, 贺彦, 郭梁, 吴建利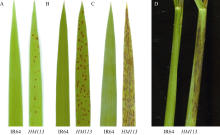










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}