关键词:大豆; 产量; 遗传进展; 遗传相关 Genetic Gain of Soybean Breeding for Yield in Henan Summer Soybean Zone over the Last 30 Years HUANG Zhong-Wen, XU Xin-Juan, WANG Wei, MEI Pei-Pei School of Life Science and Technology, Henan Institute of Science and Technology / Collaborative Innovation Center of Modern Biological Breeding of Henan Province, Xinxiang 453003, China Fund:This study was supported by the Key Technology R&D Program of Henan Province (142102110042), the Henan Provincial Project for Young Teachers in Universities (2011GGJS-133), and the Natural Science Foundation of the Education Department of Henan Province (2011A210005), and the Construction Project of Preponderant or Characteristic Discipline Crop Science in Henan Province AbstractHigh yield is a main objective in soybean breeding. This study aimed at guiding soybean breeding by understanding genetic gain of yield and its related traits. 18 soybean cultivars randomly selected from cultivars released in Henan province during the last 30 years were evaluated in two years. The results showed an increasing trend of yield with the average annual rate of 0.7%, and the genetic gain was 17.39 kg ha-1. Effective branch number, node number of main stem, seed weight, and plant height showed a positive genetic gain, whereas pod number per plant and seed number per pod showed a negative genetic gain; however, these genetic gains were weak with no significance. Soybean yield had significantly genetic and phenotypic correlations with seed weight, node number of main stem, and plant height but had no significant correlation with environment. This result indicated that seed weight, node number on main stem, and plant height had high genetic heritability and were selected simultaneously with the genetic improvement of yield, with minor effect of environment. In contrast, effective branch number, pod number per plant, and plant height had no significant correlation with yield, mainly because of their negatively genetic correlations with seed weight, node number of main stem and plant height.
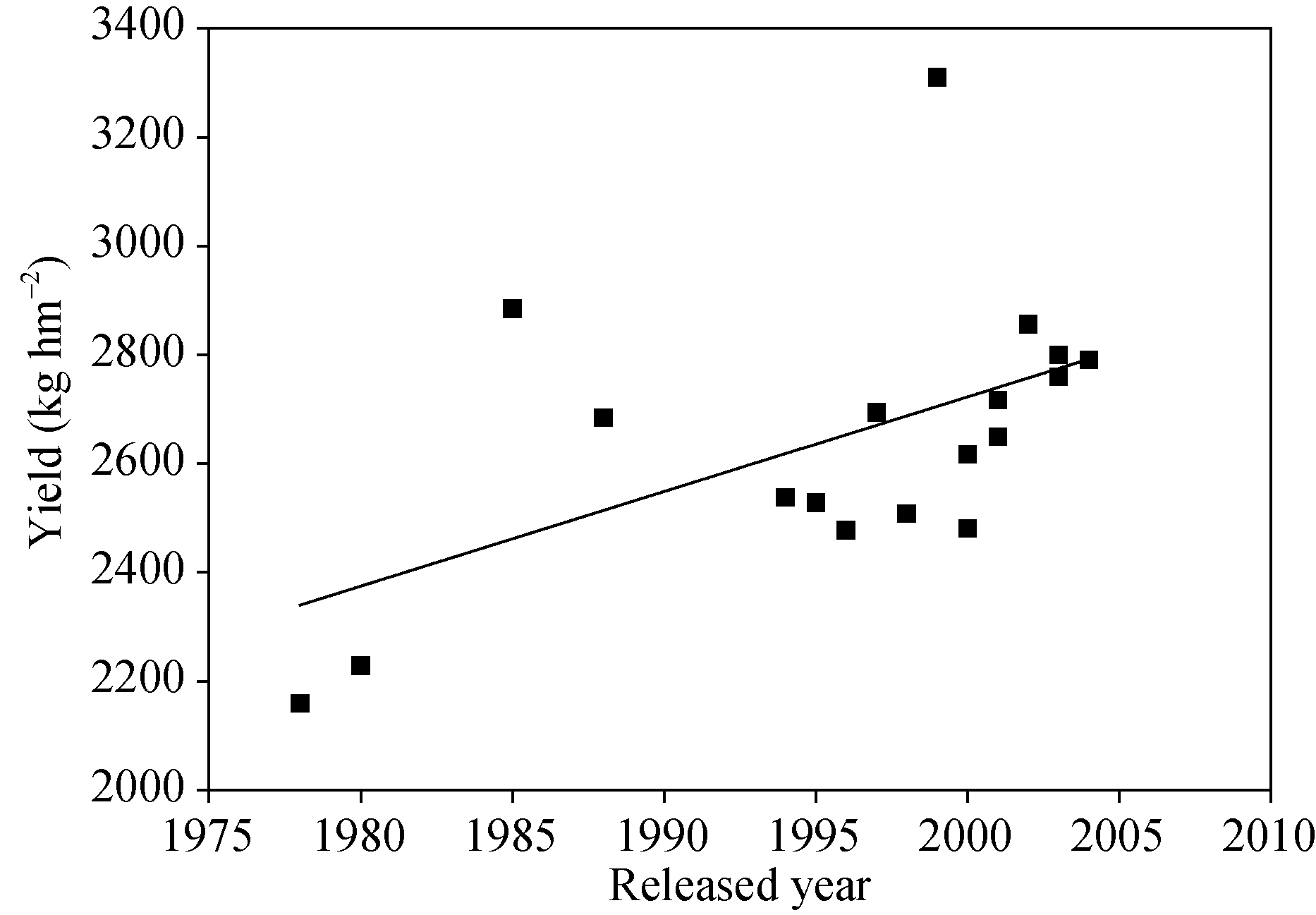
2 结果与分析2.1 产量及产量相关性状的演变2.1.1 产量 品种之间产量有极显著差异, 品种之间产量与环境的互作达到显著水平。参试品种平均产量为2641 kg hm-2, 豫豆27产量最高, 达到3310 kg hm-2, 其次是商丘1099、豫豆2号、周豆11、郑交107和郑交9525, 最低的是1978年推出的周7327-118, 仅为2160 kg hm-2 (表1)。随品种育成年份增加, 产量总体呈递增趋势, 遗传进展为17.390 kg hm-2 (R2= 0.29, P < 0.05)(图1), 年递增率是0.7% (R2= 0.33, P < 0.05)。 试验范围内, 大豆产量总趋势有两个高峰, 即1985和1988年推出的豫豆2号和豫豆8号为代表的第一个高峰, 以及以1999年推出的豫豆27为代表的第2个高峰, 30年产量变化总趋势是升高、降低、再升高。由表1和图1可见, 品种产量潜力变化可以分为3个变化阶段, 1990年以前育成的周7327-118, 商丘7608, 豫豆2号和豫豆8号为一个阶段, 其中豫豆2号产量最高, 达到2885 kg hm-2, 而周7327-118, 商丘7608和豫豆8号产量显著低于豫豆2号, 产量潜力呈增长趋势。第二阶段为1994— 1999年, 产量呈现先上升后下降再上升的趋势, 1999年育成品种豫豆27产量最高, 为3310 kg hm-2, 有效分枝也较高, 为3.3, 和豫豆21的有效分枝(3.7)差异不显著, 其株高也最高, 为107 cm, 该品种在1995— 1998年63点次试验中, 较对照豫豆8号平均增产9.5%, 其适应范围广, 对肥力要求不严, 中等以上肥力均可种植, 在河南省各地和安徽北夏大豆产区, 江苏、湖北、山东、山西、陕西等省与河南省接壤地区均可种植。第3个阶段为2000— 2004年, 2002年育成的品种商丘1099产量较高, 为2857 kg hm-2, 与之后育成的品种产量差异不显著, 品种产量潜力递增不明显。从品种产量潜力演变阶段分析, 第1和第2阶段大豆育成品种产量潜力有显著提高, 2000年后育成品种产量进展缓慢。 表1 Table 1 表1(Table 1)
表1 产量及相关性状的方差分析 Table 1 ANOVA of yield and related traits
品种 Cultivar
育成年份 Released year
产量 Yield (kg hm-2)
单株荚数 Pods per plant
每荚粒数 Seeds per pod
百粒重 100-seed weight (g)
有效分枝 Effective branch
主茎节数 Nodes on main stem
株高 Plant height (cm)
周7327-118 Zhou 7327-118
1978
2160 D
70.8 A
2.4 DBCE
16.1 IJ
2.6 ABCD
14.8 CDEF
89 CBDE
商丘7608 Shangqiu 7608
1980
2229 CD
52.5 BCDE
2.4 DBC
23.3 BC
2.8 ABCD
14.0 EF
79 CDEF
豫豆2号 Yudou 2
1985
2885 AB
65.3 ABC
2.3 DGHE
20.4 DFE
3.1 ABC
17.6 A
93 CB
豫豆8号 Yudou 8
1988
2685 BCD
67.2 AB
2.2 IGH
18.0 IFGH
2.8 ABCD
13.4 F
74 GF
豫豆16 Yudou 16
1994
2538 BCD
45.8 DE
2.4 DBC
24.0 B
2.8 ABCD
14.4 DEF
92 CBD
豫豆19 Yudou 19
1995
2528 BCD
41.2 E
2.7 A
23.6 BC
2.0 CD
16.4 ABCD
89 CBDE
豫豆21 Yudou 21
1996
2478 BCD
57.5 ABCDE
2.2 IGHE
20.4 DFE
3.7 A
14.8 BCDEF
87 CBDEF
豫豆22 Yudou 22
1997
2695 BCD
61.2 ABCD
2.2 IGH
19.0 DFEGH
3.2 AB
17.0 ABC
88 CBDEF
豫豆25 Yudou 25
1998
2508 BCD
52.4 BCDE
2.4 DBCE
19.6 DFEG
3.5 A
16.2 ABCD
88 CBDE
豫豆27 Yudou 27
1999
3310 A
63.2 ABC
2.3 DGCE
21.6 DC
3.3 AB
15.1 BCDEF
107 A
豫豆29 Yudou 29
2000
2617 BCD
69.6 A
2.1 IH
19.9 DFEG
3.2 AB
17.1 AB
77 GEF
豫豆28 Yudou 28
2000
2481 BCD
50.6 BCDE
2.3 DGHE
21.3 DCE
1.9 D
16.1 ABCDE
99 AB
郑92116 Zheng 92116
2001
2717 BCD
52.5 BCDE
2.3 DGCE
20.4 DFE
3.0 ABCD
15.5 ABCDEF
85 CBDEF
郑90007 Zheng 90007
2001
2650 BCD
71.8 A
2.4 DBCE
17.7 IGH
3.0 ABCD
16.8 ABC
85 CBDEF
商丘1099 Shangqiu 1099
2002
2857 AB
62.0 ABCD
2.5 AB
17.0 IJH
2.3 BCD
16.0 ABCDE
97 AB
周豆11 Zhoudou 11
2003
2800 ABC
48.5 CDE
2.2 IGH
27.2 A
2.5 ABCD
17.0 ABC
93 CB
郑交107 Zhengjiao 107
2003
2760 ABCD
61.8 ABCD
2.5 ABC
14.9 J
3.1 ABC
13.6 F
65 G
郑交9525 Zhengjiao 9525
2004
2791 ABC
51.4 BCDE
2.1I
18.9 FEGH
3.6 A
15.8 ABCDE
78 GDEF
平均Mean
2641
58.1
2.33
20.2
2.9
15.7
87
误差 SE
391
12.32
0.14
1.92
0.81
1.63
10.50
基因型 G
2.65* *
3.26* *
7.26* *
15.08* *
2.24*
3.57* *
4.72* *
基因× 环境 G× E
1.70*
2.40* *
2.29* *
3.24* *
1.19
0.88
1.58
遗传进展 Genetic gain
17.390*
-0.242
-0.003
0.005
0.008
0.049
0.042
决定系数 R2
0.29
0.04
0.02
0.01
0.02
0.09
0.02
Means followed by the same letter within a column are not significantly different at the 5% probability level. G and G × E mean F-value of genotype and interaction between genotype and environment, respectively. * and * * represent significance at the 0.05 and 0.01 probability levels, respectively. R2: coefficient of determination. 同一列内标以相同字母的数据表示差异不显著(P < 0.05)。G和G× E分别为基因型、基因型与环境互作的F值。* 和* * 分别表示0.05和0.01显著水平。R2为模型决定系数。
表1 产量及相关性状的方差分析 Table 1 ANOVA of yield and related traits
杨青春, 耿臻, 苑保军, 吕广伦, 张东辉. 河南省大豆生产现状及高产育种方向探讨. 大豆通报, 2000, (1): 4-6Yang QC, GengZ, Yuan BJ, Lü GL, Zhang DH. Current situation of soybean production in Henan province and the direction of high yield breeding. J Soybean Bull, 2000, (1): 4-6 (in Chinese with English abstract)[本文引用:1]
[2]
王曙明, 孟凡凡, 郑宇宏, 范旭红, 张云锋. 大豆高产育种研究进展. 中国农学通报, 2010, 26(9): 162-166Wang SM, Meng FF, Zheng YH, Fan XH, Zhang YF. Advance in research on high yield breeding of soybean. Chin Agric Sci Bull, 2010, 26(9): 162-166 (in Chinese with English abstract)[本文引用:2]
[3]
Wang BQ, Zhang LF, Dai HY, Wang CJ, LiW, XuR. Genetic variation analysis, correlation analysis and principal component analysis on agronomic traits of summer sowing soybean (Glycine max Merr. ) in Huang-Huai-Hai Region. Agric Biotechnol, 2013, 3(3): 25-29[本文引用:1]
[4]
管建慧, 郭新宇, 刘克礼. 作物品种产量潜力研究概况. 中国农学通报, 2007, 23: 202-205Guan JH, Guo XY, Liu KL. Research advances on yield potential of cultivar. Chin Agric Sci Bull, 2007, 23: 202-205 (in Chinese with English abstract)[本文引用:1]
[5]
姜国基, 刘栋锋, 张淑清. 大豆品种产量潜力的研究概况. 辽宁农业科学, 2011, (3): 52-53Jiang GJ, Liu DF, Zhang SQ. Study on yield potential of soybean cultivars. Liaoning Agric Sci, 2011, (3): 52-53 (in Chinese with English abstract)[本文引用:1]
[6]
周阳, 何中虎, 陈新民, 王德森, 张勇, 张改生. 30余年来北部麦区小麦品种产量改良遗传进展. 作物学报, 2007, 33: 1530-1535ZhouY, He ZH, Chen XM, Wang DS, ZhangY, Zhang GS. More than 30 years in northern wheat varieties yield improved genetic progress. Acta Agron Sin, 2007, 33: 1530-1535 (in Chinese with English abstract)[本文引用:1]
[7]
Johnson CB. Physiological Processes Limiting Plant Productivity. London: Butterworth Press, 1981. p 395[本文引用:1]
[8]
TeferaH, Kamara AY, AsafoadjeiB, Dashiell KE. Improvements in grain and fodder yields of early maturing promiscuous soybean cultivars in the Guinea savannas of Nigeria. Crop Sci, 2009, 49: 2037-2042[本文引用:3]
[9]
Cooper RL, MendiolaT, St-Martin S K, Fioritto R J, Dorrance A E. Registration of Apex soybean. Crop Sci, 2003, 43: 1563[本文引用:1]
[10]
Scott CR, Justin JS, Nicholas HW, Eric WW, Vince MD, Seth LN, Shaun NC, Brian WD, Paul DE, James ES, Shawn PC. Genetic gain x management interactions in soybean: I. Planting date. Crop Sci, 2013, 53: 5-7[本文引用:1]
[11]
Specht JE, Hume DJ, Kumudini SV. Soybean yield potential: a genetic and physiological perspective. Crop Sci, 1999, 39: 1560-1570[本文引用:3]
[12]
Kamara AY, MenkirA, Fakorede M A B, Ajala S O, Badu- Apraku B, Kureh I. Agronomic performance of maize cultivars representing three decades of breeding in the Guinea savannas of West and Central Africa. J Agric Sci, 2004, 142: 567-575[本文引用:2]
[13]
黄中文, 赵团结, 喻德跃, 陈受宜, 盖钧镒. 大豆产量有关性状QTL的检测. 中国农业科学, 2009, 42: 4155-4165Huang ZW, Zhao TJ, Yu DY, Chen SY, Gai JY. Detection of QTLs of yield related traits in soybean. Sci Agric Sin, 2009, 42: 4155-4165 (in Chinese with English abstract)[本文引用:1]
[14]
JinJ, Liu XB, Wang GH, MiL, Shen ZB, Chen XL, Herbert, Stephen J. Agronomic, physiological contributions to the yield improvement of soybean cultivars released from1950 to 2006 in Northeast China. Field Crops Res, 2010, 115: 116-123[本文引用:2]
[15]
李卫东. 现代河南大豆品种及栽培技术. 北京: 中国农业科学技术出版社, 2008. p 3Li WD. Modern soybean varieties and cultivation techniques in Henan. Beijing: China Agricultural Science and Technology Press, 2008. p 3[本文引用:1]
[16]
王树峰, 卢为国, 李卫东. 河南省大豆审定品种系谱来源及性状分析. 河南农业科学, 2015, 44(1): 37-41Wang SF, Lu WG, Li WD. Pedigree and trait analysis of soybean varieties registered in Henan province. J Henan Agric Sci, 2015, 44(1): 37-41 (in Chinese with English abstract)[本文引用:2]
[17]
Ortiz-MonasterioR, Sayre KD, RajaramS, McMahon M. Genetic progress in wheat yield and nitrogen use efficiency under four nitrogen rates. Crop Sci, 1997, 37: 898-904[本文引用:1]
[18]
王连铮, 叶兴国, 刘国强, 隋德志, 王培英. 黑龙江省及黄淮海地区大豆品种的遗传改进. 中国油料作物学报, 1998, 20(4): 20-25Wang LZ, Ye XG, Liu GQ, Sui DZ, Wang PY. Genetic improvement of soybean cultivars in Heilongjiang province and Huang-Huai-Hai Valley. Chin J Oil Crop Sci, 1998, 20(4): 20-25 (in Chinese with English abstract)[本文引用:2]
[19]
王振民, 康波, 邓劭华, 刘维莉, 姚唐忠. 吉林省不同年份主推大豆品种性状演变规律的初步分析. 吉林农业大学学报, 1993, (1): 92-95Wang ZM, KangB, Deng SH, Liu WL, Yao TZ. Preliminary analysis of the evolution of the main soybean cultivars in different years in Jilin province. J Jilin Agric Univ, 1993, (1): 92-95 (in Chinese with English abstract)[本文引用:1]
[20]
谢甫绨, 李契, 张惠君, 王海英, StevenK, MartinSt. 亚有限型大豆品种单株生产潜力的比较研究. 大豆科学, 2007, 26: 675-679Xie FT, LiQ, Zhang HJ, Wang HY, StevenK, MartinSt. Comparison on yield potential of individual plant between semi- determinate soybeans. Soybean Sci, 2007, 26: 675-679 (in Chinese with English abstract)[本文引用:1]
[21]
郑洪兵, 徐克章, 赵洪祥, 李大勇, 王晓慧, 张治安, 杨光宇, 杨春明, 陆静梅. 吉林省不同年代大豆品种某些株型性状的演变. 中国油料作物学报, 2006, 28: 276-281Zheng HB, Xu KZ, Zhao HX, Li DY, Wang XH, Zhang ZA, Yang GY, Yang CM, Lu JM. Study on stature evolution of soybean cultivars in Jilin province. Chin J Oil Crop Sci, 2006, 28: 276-281 (in Chinese with English abstract)[本文引用:1]
[22]
EcochardR, RavelomanantsoaY. Genetic correlations derived from full-sib relationships in soybean. Theor Appl Genet, 1982, 63: 9-15[本文引用:1]
[23]
王文斌, 孙贵荒, 孙恩玉. 辽宁省育成大豆品种蛋白质和脂肪含量的遗传改进的进展. 辽宁农业科学, 2001, (3): 35-37Wang WB, Sun GH, Sun EY. Genetic improvement of protein and fat content of soybean cultivars in Liaoning province. Liaoning Agric Sci, 2001, (3): 35-37 (in Chinese with English abstract)[本文引用:1]
[24]
Eisen EJ, Saxton AM. Genotype by environment interactions and genetic correlations involving two environmental factors. Theor Appl Genet, 1983, 67: 75-86[本文引用:1]
[25]
Alliprand ini LF, Vello NA. Heritability and correlations among traits in four way soybean crosses. Euphytica, 2004, 136: 81-91[本文引用:1]
[26]
黄中文, 赵团结, 喻德跃, 陈受宜, 盖钧镒. 大豆生物量积累、收获指数及产量间的相关与QTL分析. 作物学报, 2008, 34: 944-951Huang ZW, Zhao TJ, Yu DY, Chen SY, Gai JY. Correlation and QTL mapping of biomass accumulation, apparent harvest index, and yield in soybean. Acta Agron Sin, 2008, 34: 944-951 (in Chinese with English abstract)[本文引用:2]
[27]
刘萌娟, 李鸣雷, 石引刚. 大豆产量与若干经济性状的遗传相关与选择. 中国农学通报, 2005, 21(11): 164-166Liu MJ, Li ML, Shi YG. Genetic correlation and selection of soybean production and some economic traits. Chin Agric Sci Bull, 2005, 21(11): 164-166 (in Chinese with English abstract)[本文引用:1]
[28]
Huang ZW, Tong CF, Bo WH, Pang XM, WangZ, Xu JC, Gai JY, Wu RL. An allometric model for mapping seed development in plants. Brief Biol, 2014, 15: 562-570[本文引用:1]
[29]
邱丽娟, 王昌陵, 周国安, 陈受宜, 常汝镇. 大豆分子育种研究进展. 中国农业科学, 2007, 40: 2418-2436Qiu LJ, Wang CL, Zhou GA, Chen SY, Chang RZ. Advance in molecular breeding of soybean. Sci Agric Sin, 2007, 40: 2418-2436 (in Chinese with English abstract)[本文引用:1]
 , 徐新娟, 王伟, 梅沛沛
, 徐新娟, 王伟, 梅沛沛



{kind=link}