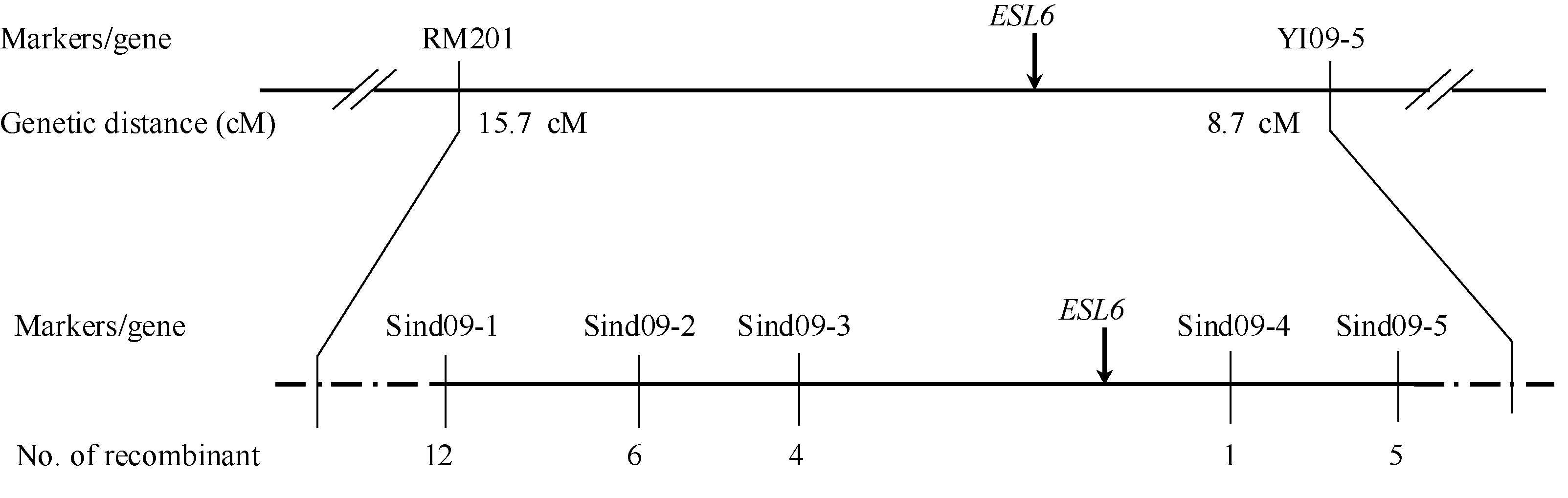
关键词:水稻( Oryza sativa L.); 早衰; 基因定位 Identification and Gene Mapping of an Early Senescent Leaf Mutant esl6 in Oryza sativa L. YANG Bo, XIA Min, ZHANG Xiao-Bo, WANG Xiao-Wen, ZHU Xiao-Yan, HE Pei-Long, HE Guang-Hua, SANG Xian-Chun* Rice Research Institute of Southwest University / Chongqing Key Laboratory of Application and Safety Control of Genetically Modified Crops, Chongqing 400715, China Fund:This study was supported by the Fundamental Research Funds for the Central Universities (XDJK2013A023) and the National Natural Science; Foundation of China (31171178) AbstractAs an essential process in life, natural senescence is necessary to adapt plant to environment diversity, while earlier senescence could reduce yield per unit and cause inferior quality in crop production. Therefore, it is significant to elucidate senescence molecular mechanism in plant. Here, we reported a novel rice mutant esl6derived from the progeny of EMS-induced restorer line Jinhui 10, which senescent peculiarity was observed at the early stage of life. In detail, cultivated under the paddy field, the esl6 had no obvious difference with the wild type before the 4-leaf stage, while after that the whole leaf blade of esl6 displayed chlorosis in the tip and kept normal green in the base until the flowering stage. Subsequently, all leaf blades in the esl6 demonstrated chlorosis and senescence, still more severe at the upper position. Observation by scanning electron microscope showed that cell structures in the senescent location of esl6 leaf blade were abnormal and filled with ruptured cell membranes, enlarged vacuoles and broken organelles such as the chloroplasts containing incomplete stroma thylakoids and excessive starch grains. Meanwhile, early senescence significantly lessened photosynthetic pigment contents and photosynthetic rate. The activities of SOD, CAT, and POD raised and the contents of O2?, H2O2, and ·OH increased in the esl6 leaf tip, and all of the differences led to the extremely significant level compared with those of the wild type. Additionally, the mutational plant showed semi-dwarfism and shorter leaf blades, the first and second internodes decreased to the extremely significant level in statistics. Genetic analysis suggested that the mutational traits were controlled by a recessive nuclear gene. The gene was finally mapped on chromosome 9 with 203 kb physical distances between Indel markers Sind09-3 and Sind09-4 on the basis of F2 generation of Xida1A/ esl6. All of these provide a foundation for ESL6 cloning and function analysis and then are beneficial to ascertaining the molecular mechanism of senescence in Oryza sativaL.
Keyword:Rice ( Oryza sativaL.); Early senescent leaf blades; Gene mapping Show Figures Show Figures
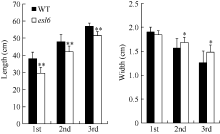
图1 突变体esl6的表型鉴定 A: 孕穗前期esl6和野生型的植株形态; B: 孕穗前期esl6和野生型的叶片; C: 灌浆期esl6和野生型的植株形态; D: 灌浆期esl6和野生型的叶片; E: 灌浆期esl6和野生型的株高构成。Fig. 1 Phenotype identification of the esl6 mutant A: Plant phenotype of the esl6 and wild type at the earlier heading stage; B: Leaf blades of the esl6 and wild type at the earlier heading stage; C: Plant phenotype of the esl6 and wild type at the filling stage; D: Leaf blades of the esl6 and wild type at the filling stage; E: Comparison of panicle and internodes between theesl6and wild type.
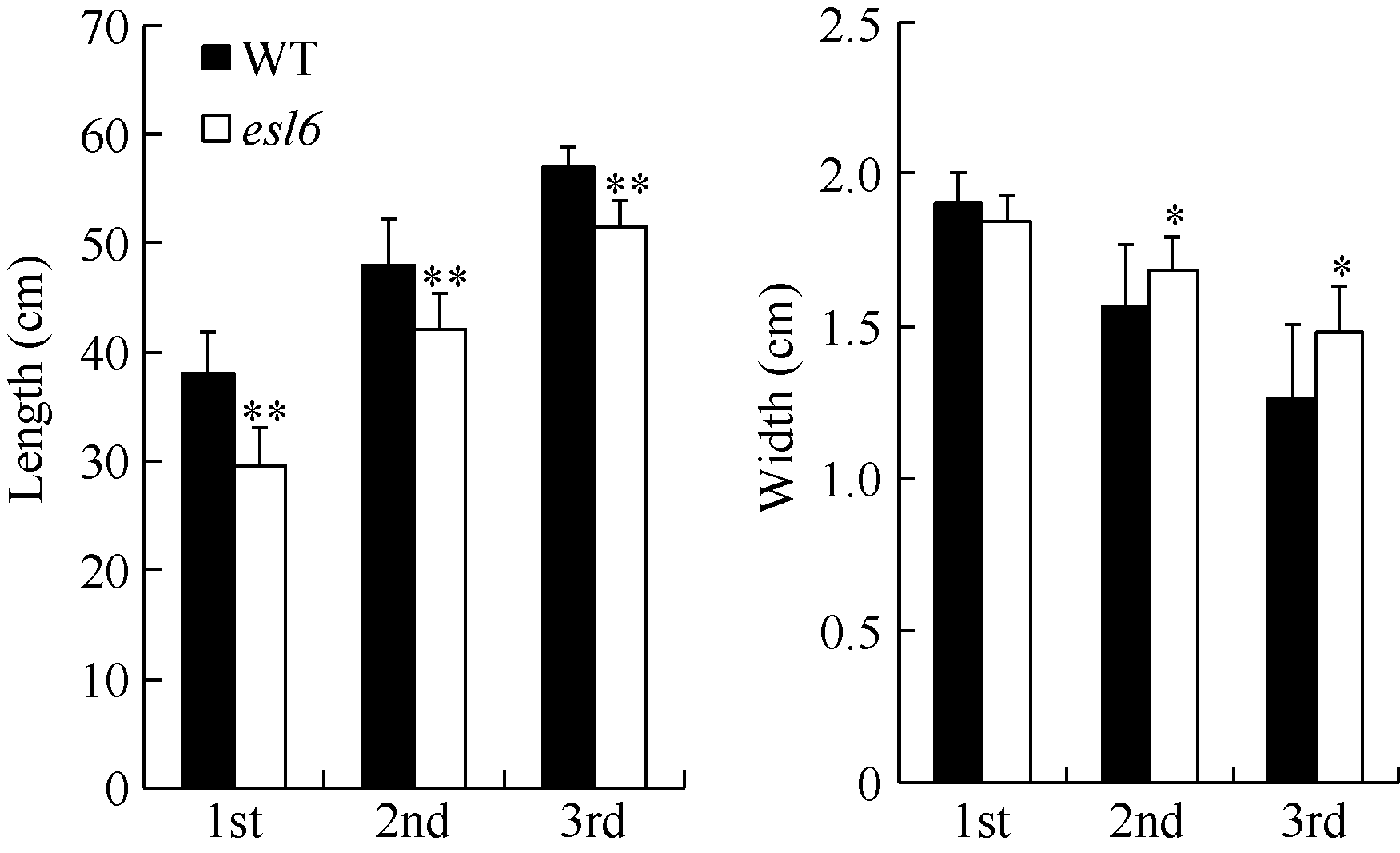
图2 灌浆期esl6的功能叶长宽分析 A: 灌浆期倒一、倒二和倒三叶的叶片长度; B: 灌浆期倒一、倒二和倒三叶的叶片宽度。Fig. 2 Leaf length and width of the esl6 and wild type during the filling stage A: Length of the first, second and third leaf blades in the wild type and esl6 at the filling stage; B: Width of the first, second and third leaf blades in the wild type and esl6 at the filling stage.
表1 Table 1 表1(Table 1)
表1 突变体esl6和野生型(WT)之间的农艺性状分析 Table 1 Agronomic analysis of the esl6 and wild type (WT)
材料 Material
有效穗数 Number of effective panicles
穗实粒数 Filled grain number per panicle
一次枝梗数 Primary branch number
二次枝梗数 Secondary branch number
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
WT
10.2± 1.34
133.20± 0.64
11.00± 1.82
53.00± 2.30
81.26± 2.14
26.54± 0.36
esl6
11.6± 1.67
85.80± 0.35* *
7.60± 0.48* *
19.20± 1.86* *
83.30± 2.45
25.71± 0.41
* * 表示在0.01水平上差异显著。* * represents the significant difference at P< 0.01.
表1 突变体esl6和野生型(WT)之间的农艺性状分析 Table 1 Agronomic analysis of the esl6 and wild type (WT)
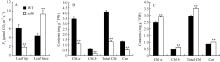
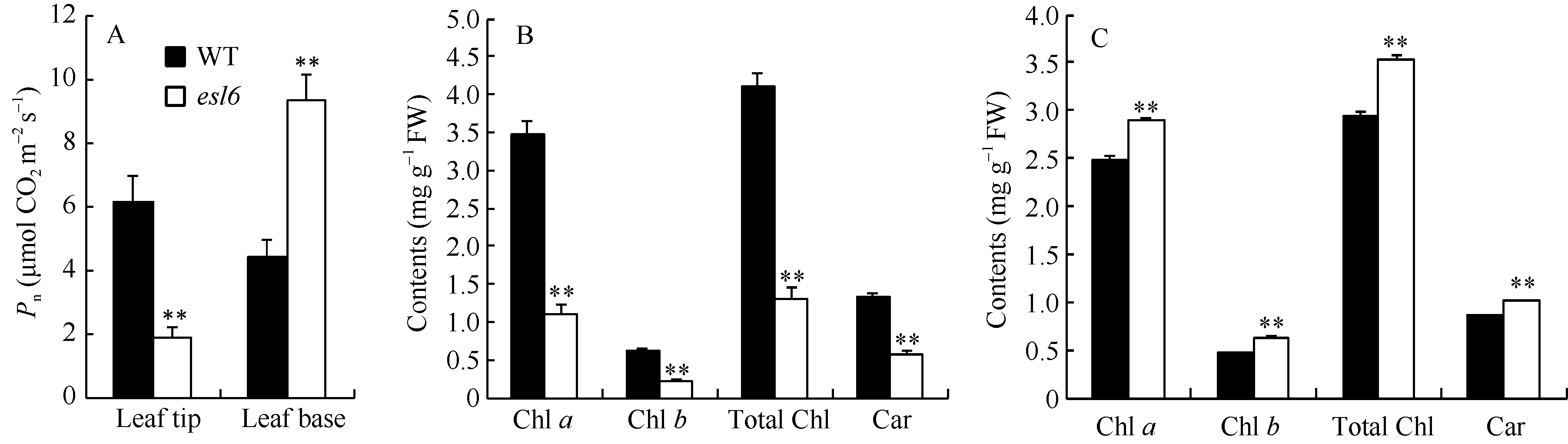
2.2 esl6净光合速率和光合色素含量孕穗期, 叶尖衰老部位, esl6的净光合速率为1.86 μ mol CO2 m-2 s-1, 极显著低于野生型的6.14 μ mol CO2 m-2 s-1; 而在叶片基部, esl6的净光合速率为9.31 μ mol CO2 m-2 s-1, 极显著高于野生型的4.42 μ mol CO2 m-2 s-1(图3-A)。光合色素含量分析发现, 叶尖衰老部位esl6的光合色素含量极显著降低(图3-B); 叶片基部, esl6的光合色素含量则极显著升高(图3-C), 与野生型相比, 叶绿素a、叶绿素b、总叶绿素和类胡萝卜素含量分别升高了16.04%、36.96%、19.32%和18.82%。光合色素含量变化与净光合速率变化趋势一致。 图3 Fig. 3
图3 孕穗期野生型(WT)和esl6的净光合速率和光合色素含量 A: 野生型和esl6叶尖和叶基部净光合速率; B: 野生型和esl6叶尖衰老部位光合色素含量; C: 野生型和esl6叶片基部持绿部位光合色素含量。Fig. 3 Photosynthetic pigment contents and net photosynthetic rate of the wild type (WT) and esl6 A: Net photosynthetic rates of the wild type and esl6 in leaf tip and leaf base; B: Photosynthetic pigment contents of the senescent leaf tips in the wild type and esl6; C: Photosynthetic pigment contents of the green leaf bases in the wild type and esl6.
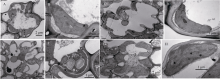
图4 突变体esl6透射电镜观察 A: 突变体esl6叶片基部细胞结构; B: 突变体esl6叶片基部叶绿体结构; C: 野生型叶片基部细胞结构; D: 野生型叶片基部叶绿体结构; E: 突变体esl6叶尖衰老部位细胞结构; F: 突变体esl6叶尖衰老部位叶绿体结构; G: 野生型叶尖部位细胞结构; H: 野生型叶尖部位叶绿体结构。Fig. 4 Cell structure of the esl6 mutant observed by the scanning electron microscope A: Cell structure in the base of esl6 leaf blade; B: Chloroplast structure in the base of esl6 leaf blade; C: Cell structure in the base of wild type leaf blade; D: Chloroplast structure in the base of wild type leaf blade; E: Cell structure of the senescent leaf tip in the esl6; F: chloroplast structure of the senescent leaf tip; G: cell structure of the wild type leaf tip; H: chloroplast structure of the wild type leaf tip.
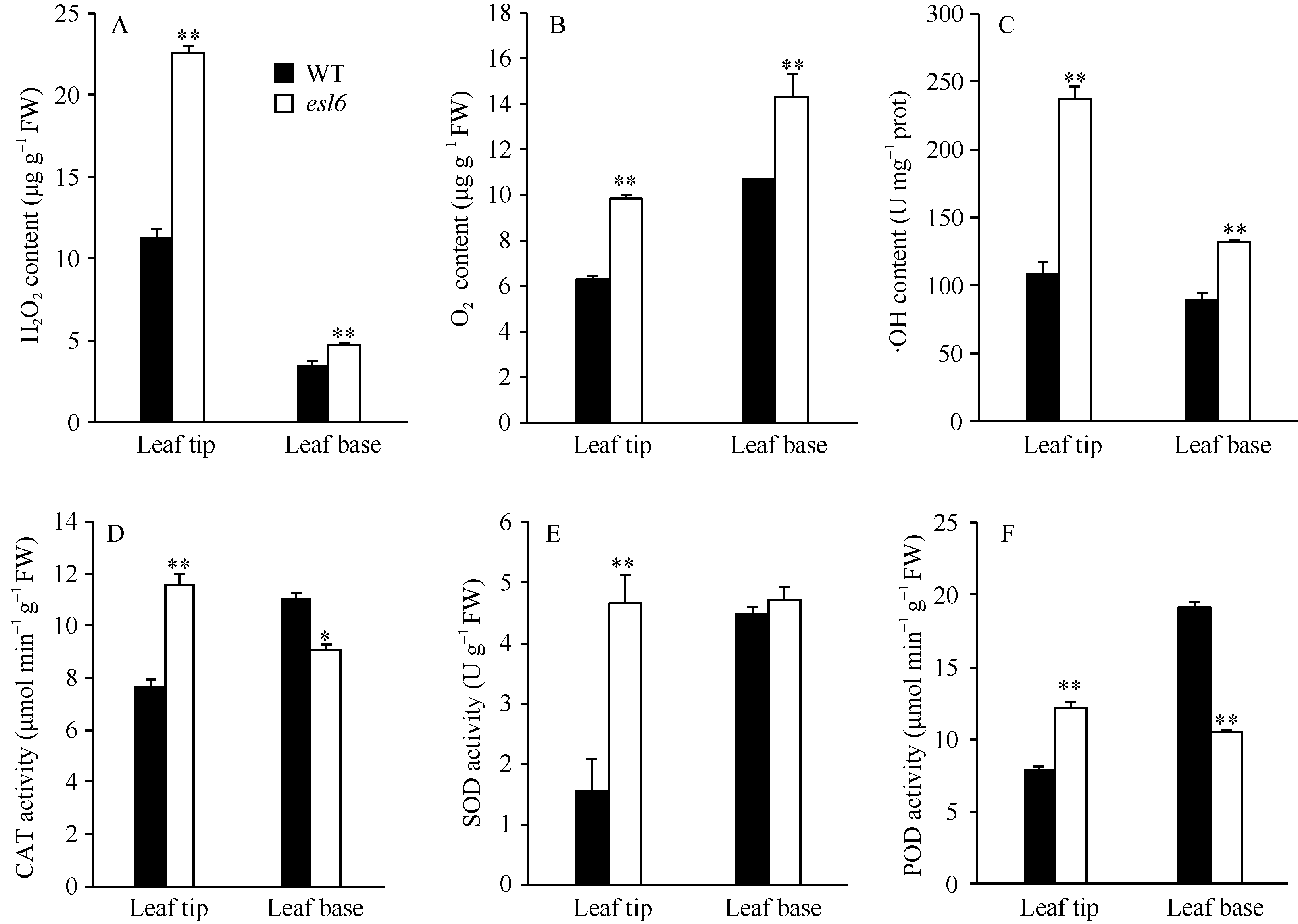
图5 抽穗期野生型(WT)和突变体esl6的生理特性 A: 野生型和突变体esl6叶尖部和叶基部过氧化氢(H2O2)含量分析; B: 野生型和esl6叶尖和叶基部超氧阴离子(O2?)分析; C: 野生型和esl6叶尖和叶基部羟自由基(· OH)分析; D: 野生型和esl6叶尖和叶基部过氧化氢酶(CAT)活性分析; E: 野生型和esl6叶尖和叶基部超氧化物歧化酶(SOD)活性分析; F: 野生型和esl6叶尖和叶基部过氧化物酶(POD)活性分析。* 和* * 分别表示在P< 0.05和P< 0.01水平上差异显著。Fig. 5 Physiological characteristics of the wild type (WT) and esl6 at the heading stage A: H2O2 contents in leaf tip and leaf base; B: Content of superoxide anion (O2?) in leaf tip and leaf base; C: Content of hydroxyl radical (· OH) in leaf tip and leaf base; D: Catalase (CAT) activity in leaf tip and leaf base; E: Activity of superoxide dismutase (SOD) in leaf tip and leaf base; F: Activity of peroxidase (POD) in leaf tip and leaf base. * and * * represent the significant difference at P< 0.05 and P< 0.01, respectively.
Kim HJ, Lim PO, Hong GN. Molecular regulation of leaf senescence. Curr Opin Plant Biol, 2003, 6: 79-84[本文引用:1]
[2]
LeisterD. Chloroplast research in the genomic age. Trends Genet, 2003, 19: 47-56[本文引用:1]
[3]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
LiuL, ZhouY, ZhouG, Ye RJ, Zhao LN, Li XH, Lin YJ. Identification of early senescence-associated genes in rice flag. Plant Mol Biol, 2008, 67: 37-55[本文引用:1]
[6]
Li ZH, ZhaoY, Liu XC, Peng JY, Guo HW, Luo JC. LSD 2. 0: an update of the leaf senescence database. Nucl Acids Res, 2014, 42(D1): 200-205[本文引用:1]
[7]
Xu XB, Bai HQ, Liu CP, Chen EY, Chen QF, Zhuang JY, ShenB. Genome-wide analysis of microRNAs and their target genes related to leaf senescence of rice. PLoS One, 2014, 9(12): e114313[本文引用:1]
[8]
Schippers J H M, Schmidt R, Wagstaff C, Jing H C. Living to die and dying to live: The survival strategy behind leaf senescence. Plant Physiol, 2015, 169: 914-930[本文引用:1]
[9]
Abdelkhalik AF, Nomura R S K, Ikehashi H. QTL-based analysis of leaf senescence in an indica/japonica hybrid in rice (Oryza sativa L. ). Theor Appl Genet, 2005, 110: 1226-1235[本文引用:1]
[10]
Yoo SC, Cho SH, ZhangH, Paik HC, Lee CH, LiJ, Yoo JH, Lee BW, Koh HJ, Seo HS, Paek NC. Quantitative trait loci associated with functional stay-green SNU-SG1 in rice. Mol Cells, 2007, 24: 83-94[本文引用:1]
[11]
Wu HB, WangB, Chen LY, Liu LG, Chen LT. Characterization and fine mapping of the rice premature senescence mutant ospse1. Theor Appl Genet, 2013, 126: 1897-1907[本文引用:1]
[12]
LiZ, Zhang YX, LiuL, LiuQ, Bi ZZ, YuN, Cheng SH, Cao LY. Fine mapping of the lesion mimic and early senescence 1 (lmes1) in rice (Oryza sativa). Plant Physiol Biochem, 2014, 80: 300-307[本文引用:1]
[13]
Yan WY, Ye SH, Jin QS, Zeng LJ, PengY, Yan DW, Yang WB, Yang DL, He ZH, Dong YJ, Zhang XM. Characterization and mapping of a novel mutant sms1 (senescence and male sterility 1) in rice. J Genet Genomics, 2010, 37: 47-55[本文引用:1]
[14]
WangJ, Wu SJ, ZhouY, Zhou LH, Xu JF, HuJ, Fang YX, Gu MH, Liang GH. Genetic analysis and molecular mapping of a presenescing leaf gene psl1 in rice (Oryza sativa L. ). Chin Sci Bull, 2006, 51: 2986-2992[本文引用:1]
[15]
张涛, 孙玉莹, 郑建敏, 程治军, 蒋开锋, 杨莉, 曹应江, 游书梅, 万建民, 张建奎. 水稻早衰叶突变体PLS2的遗传分析与基因定位. 作物学报, 2014, 40: 2070-2080ZhangT, Sun YY, Zheng JM, Cheng ZJ, Jiang KF, YangL, Cao YJ, You SM, Wan JM, Zhang JK. Genetic analysis and fine mapping of a premature leaf senescence mutant in rice (Orzya sativa L. ). Acta Agron Sin, 2014, 40: 2070-2080 (in Chinese with an English abstract)[本文引用:1]
[16]
Li FZ, Hu GC, Fu YP, Si HM, Bai XM, Sun ZX. Genetic analysis and high-resolution mapping of a premature senescence gene Pse(t) in rice (Oryza sativa L. ). Genome, 2005, 48: 738-746[本文引用:1]
[17]
Fang LK, Li YF, Gong XP, Sang XC, Ling YH, Wang XW, Cong YF, He GH. Genetic analysis and gene mapping of a dominant presenescing leaf gene PSL3 in rice (Oryza sativa L. ). Chin Sci Bull, 2010, 55: 2517-2521[本文引用:2]
[18]
Yang YL, Rao YC, Liu HJ, Fang YX, Dong GJ, Huang LC, Leng YJ, Guo LB, Zhang GH, HuJ. Characterization and fine mapping of an early senescence mutant (es-t) in Oryza sativa L. Chin Sci Bull, 2011, 56: 2437-2443[本文引用:1]
[19]
徐芳芳, 桑贤春, 任德勇, 唐彦强, 胡宏伟, 杨正林, 赵芳明, 何光华. 水稻早衰突变体esl2的遗传分析和基因定位. 作物学报, 2012, 38: 1347-1353Xu FF, Sang XC, Ren DY, Tang YQ, Hu HW, Yang ZL, Zhao FM, He GH. Genetic analysis and gene mapping of early senescence leaf mutant esl2 in rice. Acta Agron Sin, 2012, 38: 1347-1353 (in Chinese with an English abstract)[本文引用:1]
[20]
吕典华, 宗学凤, 王三根, 凌英华, 桑贤春, 何光华. 两个水稻叶色突变体的光合特性研究. 作物学报, 2009, 35: 2304-2308. Lv DH, Zong XF, Wang SG, Ling YH, Sang XC, He GH. Characteristics of photosynthesis in two leaf color mutants of rice. Acta Agron Sin, 2009, 35: 2304-2308 (in Chinese with an English abstract)[本文引用:1]
[21]
Wellburn AR. The spectra determination of chlorophylls a and b, as well as total carotenoids, using various solvents with spectrophotometers of different resolution. J Plant Physiol, 1994, 144: 307-313[本文引用:1]
[22]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1]
[23]
王晓雯, 蒋钰东, 廖红香, 杨波, 邹帅宇, 朱小燕, 何光华, 桑贤春. 水稻白穗突变体wp4的鉴定与基因精细定位. 作物学报, 2015, 41: 838-844Wang XW, Jiang YD, Liao HX, YangB, Zou SY, Zhu XY, He GH, Sang XC. Identification and gene fine mapping of white panicle mutant wp4 in Oryza sativa. Acta Agron Sin, 2015, 6: 838-844 (in Chinese with an English abstract)[本文引用:1]
[24]
PanaudO, ChenX, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSLP) in rice (Oryza sativa L. ). Mol General Genet, 1996, 252: 597-607[本文引用:1]
[25]
SulpiceR, Pyl ET, IshiharaH, TrenkampS, SteinfathM, Witucka-WallH, GibonY, UsadelB, PoreeF, Piques MC, Korff MV, Steinhauser MC, Keurentjes J J B, Guenther M, Hoehne M, Selbig J, Fernie A R, Altmann T, Stitt M. Starch as a major integrator in the regulation of plant growth. Proc Natl Acad Sci USA, 2009, 106: 10348-10353[本文引用:1]
[26]
SamojednyD, OrzechowskiS. New look at starch degradation in Arabidopsis thaliana L. chloroplasts. Postepy Biochem, 2007, 53: 74-83[本文引用:1]
[27]
Yand eau-Nelson M D, Laurens L, Shi Z, Xia H, Smith A M, Guiltinan M J. Starch-branching enzyme IIa is required for proper diurnal cycling of starch in leaves of maize. Plant Physiol, 2011, 156: 479-490[本文引用:1]
[28]
Lin AH, Wang YQ, Tang JY, XueP, Li CL, Liu LC, HuB, Yang FQ, Loake GJ, Chu CC. Nitric oxide and protein S-Nitrosylation are integral to hydrogen peroxide-induced leaf cell death in rice. Plant Physiol, 2012, 158: 451-464[本文引用:1]
[29]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1]
[30]
Kong ZS, Li MN, Yang WQ, Xu WY, Xue YB. A novel nuclear-localized CCCH-type zinc finger protein, OsDOS, is involved in delaying leaf senescence in rice. Plant Physiol, 2006, 141: 1376-1388[本文引用:1]
[31]
Liang CZ, Wang YQ, Zhu YN, Tang JY, HuB, Liu LC, Ou SJ, Wu HK, Sun XH, Chu JF. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc Natl Acad Sci USA, 2014, 111: 10013-10018[本文引用:1]
[32]
SakurabaY, PiaoW, Lim JH, Han SH, Kim YS, AnG, Paek NC. Rice ONAC10inhibits leaf senescence and increases salt tolerance and tiller angle. Plant Cell Physiol, 2015, 56: 2325-2339[本文引用:1]
[33]
Schippers J H M. Transcriptional networks in leaf senescence. Curr Opin Plant Biol, 2015, 27: 77-83[本文引用:1]
[34]
Sun SJ, Guo SQ, YangX, Bao YM, Tang HJ, SunH, HuangJ, Zhang HS. Functional analysis of a novel Cys2/His2-type zinc finger protein involved in salt tolerance in rice. J Exp Bot, 2010, 61: 2807-2818[本文引用:1]
[35]
Hu XM, QianQ, XuT, ZhangY, Dong GJ, GaoT, XieQ, Xue YB. The U-Box E3 ubiquitin ligase TUD1 functions with a heterotrimeric G α subunit to regulate Brassinosteroid-mediated growth in rice. PLoS Genet, 2013, 9(3): e1003391[本文引用:1]
[36]
Fang HM, Meng QL, Xu JW, Tang HJ, Tang SY, Zhang HS, HuangJ. Knock-down of stress inducible OsSRFP1 encoding an E3 ubiquitin ligase with transcriptional activation activity confers abiotic stress tolerance through enhancing antioxidant protection in rice. Plant Mol Biol, 2015, 87: 441-458[本文引用:1]
[37]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1]
[38]
LepistöA, KangasjärviS, Luomala EM, BraderG, SipariN, KeränenM, KeinänenM, RintamäkiE. Chloroplast NADPH- thioredoxin reductase interacts with photoperiodic development in Arabidopsis. Plant Physiol, 2009, 149: 1261-1276[本文引用:1]
 , 夏敏, 张孝波, 王晓雯, 朱小燕, 何沛龙, 何光华, 桑贤春
, 夏敏, 张孝波, 王晓雯, 朱小燕, 何沛龙, 何光华, 桑贤春





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}