关键词:大豆; GmNAC; 进化树; NaCl处理; 表达分析; 序列变异 Identification, Expression and Variation Analysis of Salt Tolerance Related GmNAC Genes in Soybean ZHANG Yan-Wei1,2, ZHANG Li-Feng1, LI Wei1, WANG Cai-Jie1, ZHANG Jun1, XU Ran1,* 1 Crop Research Institute, Shandong Academy of Agricultural Sciences, Jinan 250131, China
2 Key Laboratory of Soybean Biology of Ministry of Education, Northeast Agricultural University, Harbin 150001, China
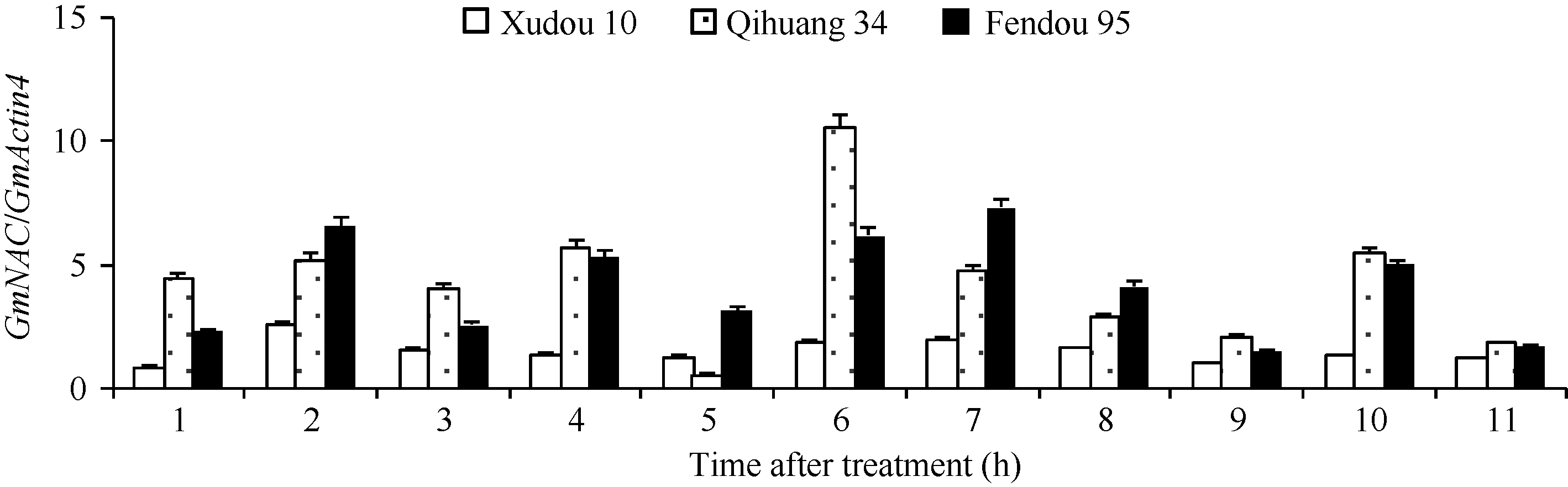
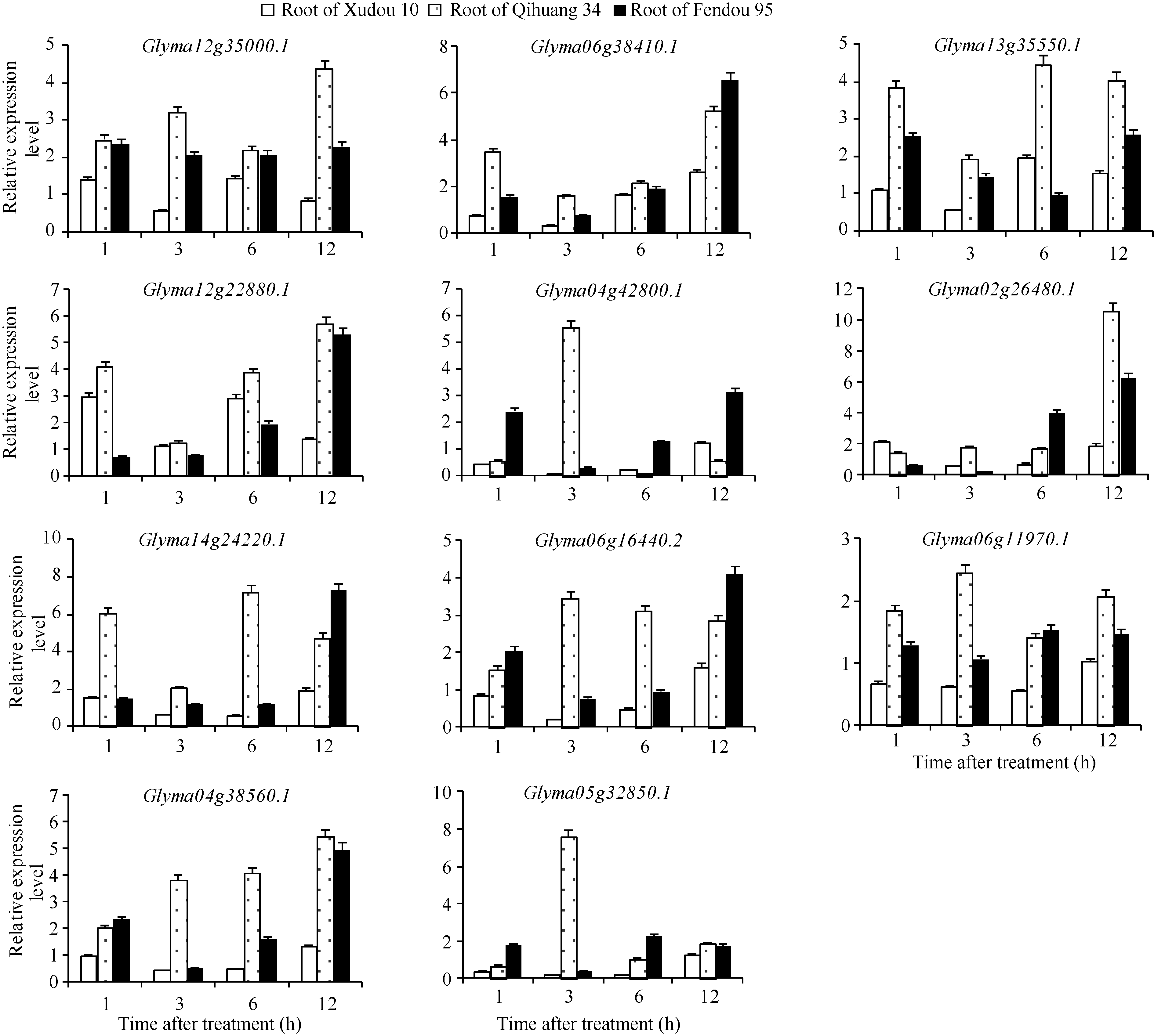
Fund:The study was supported by the Open Foundation of Key Laboratory of Soybean Biology of Ministry of Education, Northeast Agricultural University (SB14A04), the China Agriculture Research System (CARS-04-CES18), the National Natural Science Foundation of China (31501329), and the Natural Science Foundation of Shandong Province (ZR2015YL070) Abstract NAC genes play an important role in plant stress tolerance. In this study, bioinformatic method was used to identify the stress related GmNAC gene in soybean. The expression of candidated GmNAC genes in root and leaf was analyzed in soybean with NaCl treatment by Real-time PCR. Reverse transcription PCR was performed to clone genes with significant difference in expression. The results showed that there were 175 genes in soybean GmNAC gene family. There were 11 GmNAC proteins with highly conserved NAC located on the same evolutionary branch with the stress related NAC proteins in rice and Arabidopsis. The expression of 11 GmNAC genes in soybean root was higher than that in leaf. The GmNAC genes were all induced by NaCl stress, but part of the GmNAC genes showed different expression levels between root and leaf in soybean varieties with different salt tolerances. There were three synonymous mutations and one non-synonymous mutation on the CDS region of Glyma06g11970.1 and one synonymous mutation on the CDS region of Glyma06g16440.2 in Qihuang 34, Xudou 10, and Fendou 95.
Keyword:Soybean; GmNAC; Phylogenetic tree; NaCl treatment; Expression analysis; Sequence variation Show Figures Show Figures
表3 水稻和拟南芥逆境相关NAC基因 Table 3 Stress related NAC genes in rice and Arabidopsis
基因登录号 Gene ID
基因名称 Gene name
基因功能 Gene function
AT1G52890
AtNAC019
ABA信号通路; 参与ABA、热、高盐、JA、干旱应答; 生长发育[6, 7, 8]。 ABA signaling pathway; induced by ABA, heat, high salt, JA, drought; regulate plant growth and development.
AT3G15500
AtNAC055
JA生物合成, JA通路, 高盐应答, 菌防卫应答, 器官发育, 调控细胞程序性死亡, 调控防御应答[7, 8]。 JA biosynthesis; JA signaling pathway; response to high salt, fungi; organism development; negative regulation of programmed cell death; negative regulation of defense responses.
AT4G27410
AtNAC072
ABA信号通路, 参与高盐、ABA、生长素、乙烯、JA、脱水、损伤应答, 信号传导[5]。 ABA signaling pathway; induced by ABA, IAA, ethylene, JA, high salt, dehydration, wounding; signal transduction.
AT5G63790
AtNAC102
洪涝等逆境胁迫反应[9]。 Involving in response to stress such as flooding.
DQ394702.1
OsNAC002(SNAC1)
耐寒, 耐盐, 耐氧化胁迫[10, 11, 12, 13]。Involving in response to cold, high salt, oxidative stress.
AB028185.1
OsNAC048(OsNAC6)
生物和非生物胁迫应答[14, 15]。Response to biotic and abiotic stress.
AB028183.1
OsNAC068(OsNAC4)
耐旱, 耐盐[16]。Involving in response to drought, high salt.
表3 水稻和拟南芥逆境相关NAC基因 Table 3 Stress related NAC genes in rice and Arabidopsis
图1 NAC蛋白系统进化树 ●: 大豆干旱胁迫诱导的GmNAC蛋白; ○: 水稻和拟南芥中逆境相关的NAC蛋白。Fig. 1 Phylogenetic tree of NAC proteins ●: GmNAC proteins induced by drought in soybean; ○: Stress related NAC proteins in rice and Arabidopsis.
图7GmNAC基因在大豆品种中的序列变异 a: Glyma06g11970.1在大豆品种中的序列变异; b: Glyma06g16440.2在大豆品种中的序列变异。Fig. 7 Sequence variations of GmNAC in soybean varieties a: Variations of Glyma06g11970.1 in soybean varieties; b: Variations of Glyma06g16440.2 in soybean varieties.
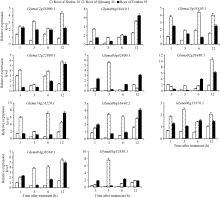
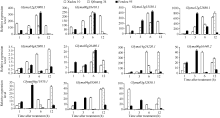
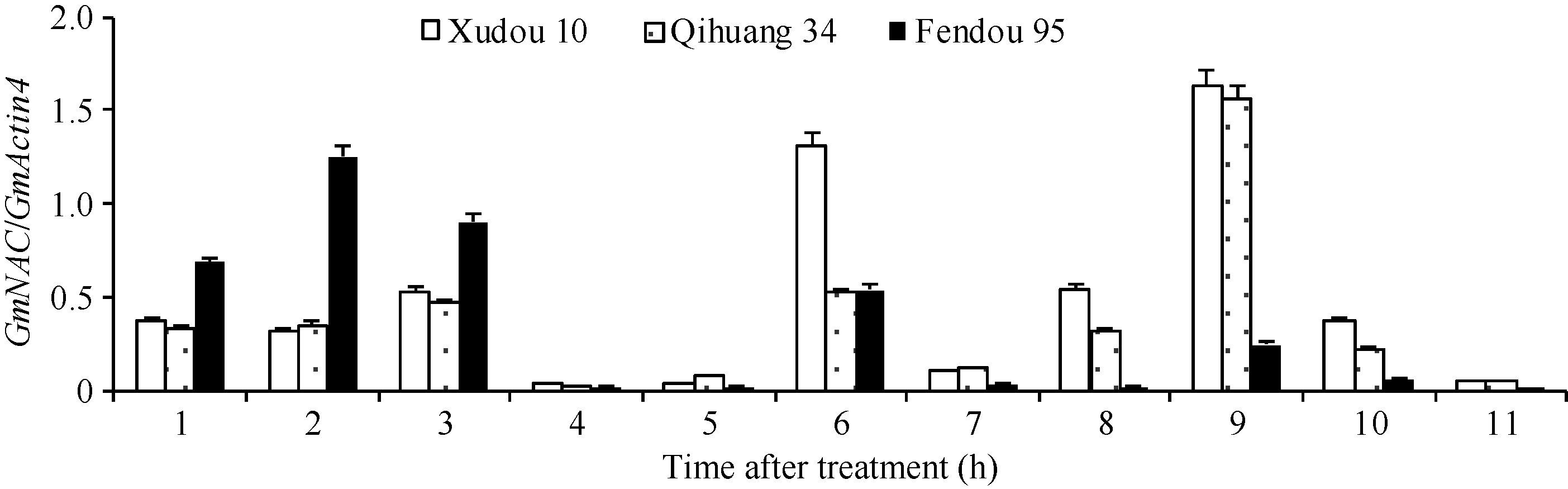
4 结论获得了3个盐胁迫相关的GmNAC基因(Glyma04g38560.1、Glyma06g11970.1和Glyma0 6g16440.2), 其中Glyma06g11970.1和Glyma0 6g16440.2在徐豆10、齐黄34和汾豆95中存在SNP位点。 The authors have declared that no competing interests exist.
Atkinson NJ, Urwin PE. The interaction of plant biotic and abiotic stresses: from genes to the field. J Exp Bot, 2012, 63: 3523-3543[本文引用:1]
[2]
李鹏, 黄耿青, 李学宝. 植物NAC转录因子. 植物生理学通讯, 2010, 46: 294-300LiP, Huang GQ, Li XB. Plant NAC transcription factors. Plant Physiol Mol Biol, 2010, 46: 294-300 (in Chinese with English abstract)[本文引用:1]
[3]
AidaM, IshidaT, FukakiH, FujisawaH, TasakaM. Genes involved in organ separation in Arabidopsis: an analysis of the cup-shaped cotyledon mutant. Plant Cell, 1997, 9: 841-857[本文引用:1]
[4]
柳展基, 邵凤霞, 唐桂英. 植物NAC转录因子的结构功能及其表达调控研究进展. 西北植物学报, 2007, 27: 1915-1920Liu ZJ, Shao FX, Tang GY. The research progress of structure, function and regulation of plant NAC transcription factors. Acta Bot Boreal-Occident Sin, 2007, 27: 1915-1920 (in Chinese with English abstract)[本文引用:1]
[5]
HickmanR, HillC, PenfoldCA, BreezeE, BowdenL, Moore JD, ZhangP, JacksonA, CookeE, Bewicke-CopleyF, MeadA, BeynonJ, Wild DL, Denby KJ, OttS, Buchanan-WollastonV. A local regulatory network around three NAC transcription factors in stress responses and senescence in Arabidopsis leaves. Plant J, 2013, 75: 26-39[本文引用:2]
[6]
GuanQ, YueX, ZengH, ZhuJ. The protein phosphatase RCF2 and its interacting partner NAC019 are critical for heat stress- responsive gene regulation and thermo tolerance in Arabidopsis. Plant Cell, 2014, 26: 438-453[本文引用:2]
[7]
BuQ, JiangH, Li CB, ZhaiQ, ZhangJ, WuX, SunJ, XieQ, LiC. Role of the Arabidopsis thaliana NAC transcription factors ANAC019 and ANAC055 in regulating jasmonic acid-signaled defense responses. Cell Res, 2008, 18: 756-767[本文引用:3]
[8]
JiangH, LiH, BuQ, LiC. The RHA2a-interacting proteins ANAC019 and ANAC055 may play a dual role in regulating ABA response and jasmonate response. Plant Signal Behav, 2009, 4: 464-466[本文引用:3]
[9]
Christianson JA, Wilson IW, Llewellyn DJ, Dennis ES. The low-oxygen-induced NAC domain transcription factor ANAC10affects viability of Arabidopsis seeds following low-oxygen treatment. Plant Physiol, 2009, 149: 1724-1738[本文引用:1]
[10]
YouJ, ZongW, LiX, NingJ, HuH, LiX, XiaoJ, XiongL. The SNAC1-targeted gene OsSRO1c modulates stomatal closure and oxidative stress tolerance by regulating hydrogen peroxide in rice. J Exp Bot, 2013, 64: 569-583[本文引用:1]
[11]
HuH, DaiM, YaoJ, XiaoB, LiX, ZhangQ, XiongL. Overexpressing a NAM, ATAF, and CUC (NAC) transcription factor enhances drought resistance and salt tolerance in rice. Proc Natl Acad Sci USA, 2006, 103: 12987-12992[本文引用:1]
[12]
KikuchiK, Ueguchi-TanakaM, Yoshida KT, NagatoY, MatsusokaM, Hirano HY. Molecular analysis of the NAC gene family in rice. Mol Gen Genet, 2000, 262: 1047-1051[本文引用:1]
[13]
LiuG, LiX, JinS, LiuX, ZhuL, NieY, ZhangX. Overexpression of rice NAC gene SNAC1 improves drought and salt tolerance by enhancing root development and reducing transpiration rate in transgenic cotton. PLoS One, 2014, 9(1): e86895[本文引用:1]
[14]
TagaY, TakaiR, KanedaT, MatsuiH, IsogaiA, Che FS. Role of OsHSP90 and IREN, Ca2+ dependent nuclease, in plant hypersensitive cell death induced by transcription factor OsNAC4. Plant Signal Behav, 2009, 4: 740-742[本文引用:1]
[15]
KanedaT, TagaY, TakaiR, IwanoM, MatsuiH, TakayamaS, IsogaiA, Che FS. The transcription factor OsNAC4 is a key positive regulator of plant hypersensitive cell death. EMBO J, 2009, 28: 926-936[本文引用:1]
[16]
NakashimaK, TranLS, Van NguyenD, FujitaM, MaruyamaK, TodakaD, ItoY, HayashiN, ShinozakiK, Yamaguchi-ShinozakiK. Functional analysis of a NAC-type transcription factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice. Plant J, 2007, 51: 617-630[本文引用:1]
[17]
孟庆长. 大豆GmNAC和GmLFY转录因子编码基因的克隆、鉴定和种子性状的QTL定位研究. 南京农业大学博士学位论文, 江苏南京, 2006Meng QC. Cloning and Identification of Genes Encoding Two Types of Transcription Factor, GmNAC and GmLFY and QTL Mapping for Seed Traits in Soybean (Glycine max (L. ) Merr. ). PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2006 (in Chinese with English abstract)[本文引用:1]
[18]
MengQ, ZhangC, GaiJ, YuD. Molecular cloning, sequence characterization and tissue-specific expression of six NAC-like genes in soybean (Glycine max (L. ) Merr. ). J Plant Physiol, 2007, 164: 1002-1012[本文引用:1]
[19]
韩巧玲. 大豆重要抗逆相关基因GmE2、GmNAC2a的特性分析及功能鉴定. 西北农林科技大学硕士学位论文, 陕西杨凌, 2010Han QL. Characteristics and Functional Identification of Two Important Stress-Related Genes GmE2 and GmNAC2a from Soybean. MS Thesis of Northwest A&F University, Yangling, China, 2010 (in Chinese with English abstract)[本文引用:1]
[20]
金杭霞. 大豆转录因子GmNAC2和GmNAC5功能验证. 南京农业大学博士学位论文, 江苏南京, 2011Jin HX. Functional Analysis of Two Transfactors GmNAC2 and GmNAC5 in Soybean. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2011 (in Chinese with English abstract)[本文引用:1]
[21]
才华, 朱延明, 李勇, 柏锡, 纪巍, 王冬冬, 孙晓丽. 野生大豆转录因子GsNAC20基因的分离及胁迫耐性分析. 作物学报, 2011, 37: 1351-1359CaiH, Zhu YM, LiY, BaiX, JiW, Wang DD, Sun XL. Isolation and tolerance analysis of GsNAC20 gene linked to response to stress inGlycine soja. Acta Agron Sin, 2011, 37: 1351-1359 (in Chinese with English abstract)[本文引用:1]
Le DT, NishiyamaR, WatanabeY, MochidaK, Yamaguchi- ShinozakiK, ShinozakiK, Tran LS. Genome-wide survey and expression analysis of the plant-specific NAC transcription factor family in soybean during development and dehydration stress. DNA Res, 2011, 18: 263-276[本文引用:4]
[24]
Le DT, NishiyamaR, WatanabeY, TanakaM, SekiM, Ham le H, Yamaguchi-Shinozaki K, Shinozaki K, Tran L S. Differential gene expression in soybean leaf tissues at late developmental stages under drought stress revealed by genome-wide transcriptome analysis. PLoS One, 2012, 7(11): e49522[本文引用:4]
[25]
王洋, 柏锡. 大豆NAC基因家族生物信息学分析. 大豆科学, 2014, 33: 325-333WangY, BaiX. Bioinformatics analysis of NAC gene family in Glycine max L. Soybean Sci, 2014, 33: 325-333 (in Chinese with English abstract)[本文引用:3]
[26]
Hao YJ, Song QX, Chen HW, Zou HF, WeiW, Kang XS, MaB, Zhang WK, Zhang JS, Chen SY. Plant NAC-type transcription factor proteins contain a NARD domain for repression of transcriptional activation. Planta, 2010, 232: 1033-1043[本文引用:4]
李伟, 韩蕾, 钱永强, 巨关升, 孙振元. 非生物逆境胁迫相关NAC转录因子的生物信息学分析. 西北植物学报, 2012, 32: 454-464LiW, HanL, Qian YQ, Ju GS, Sun ZY. Bioinformatics analysis of abiotic stress related NAC transcription factors. Acta Bot Boreal-Occident Sin, 2012, 32: 454-464 (in Chinese with English abstract)[本文引用:1]
[29]
YouJ, ZhangL, SongB, QiX, ChanZ. Systematic analysis and identification of stress-responsive genes of the NAC gene family in Brachypodium distachyon. PloS One, 2015, 10(3): e0122027[本文引用:1]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}