关键词:水稻; 基因编辑; CRISPR/Cas9; TGW6; 千粒重 Construction of tgw6 Mutants in Rice Based on CRISPR/Cas9 Technology WANG Jia-Feng, ZHENG Cai-Min, LIU Wei, LUO Wen-Long, WANG Hui, CHEN Zhi-Qiang*, GUO Tao* National Engineering Research Center of Plant Space Breeding, South China Agricultural University, Guangzhou 510642, China Fund:This study was supported by Public Welfare Research and Capacity Building Transformation Funds in Guangdong (20150209), the National High Technology Research and Development Program of China (863 Program) (2011AA10A101), and the China Agricultural Research System (CARS-01-12) AbstractA set of tgw6 ( Thousand-grain weight 6) mutants were constructed using CRISPR/Cas9 technology in this study. Three sites of 20 nt guide RNA (gRNA) targeted to the exon of TGW6were designed and transcribed from the U3, U6a, or U6b promoters, respectively. The three target sites of gRNA were then ligated to the vector pYLCRISPR/Cas9-MT(I) based on golden gate cloning strategy. The recombinant plasmid was transferred to a rice cultivar, H447 (R819/Yuzhenxiang//R819 BC3F6) by Agrobacterium-mediated transformation. Sequencing for the genomic DNA of TGW6locus in T0 rice showed the mutagenesis frequency for TGW6was more than 90%, including 51% of homozygous deletion mutations. Further analysis for the T1 mutants showed that almost all the homozygous deletion mutants improved the thousand-grain weight significantly (more than 5%). The successful tgw6 editing not only provided a series of tgw6mutants for high and stable yield of rice but also proved that CRISPR/Cas9 is a facile and powerful means of rice genetic engineering for scientific and agricultural applications, which has important theoretical and practical significance for rice breeding.
Keyword:Rice; Genome editing; CRISPR/Cas9; TGW6; Thousand-grain weight Show Figures Show Figures
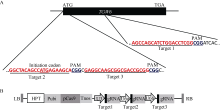
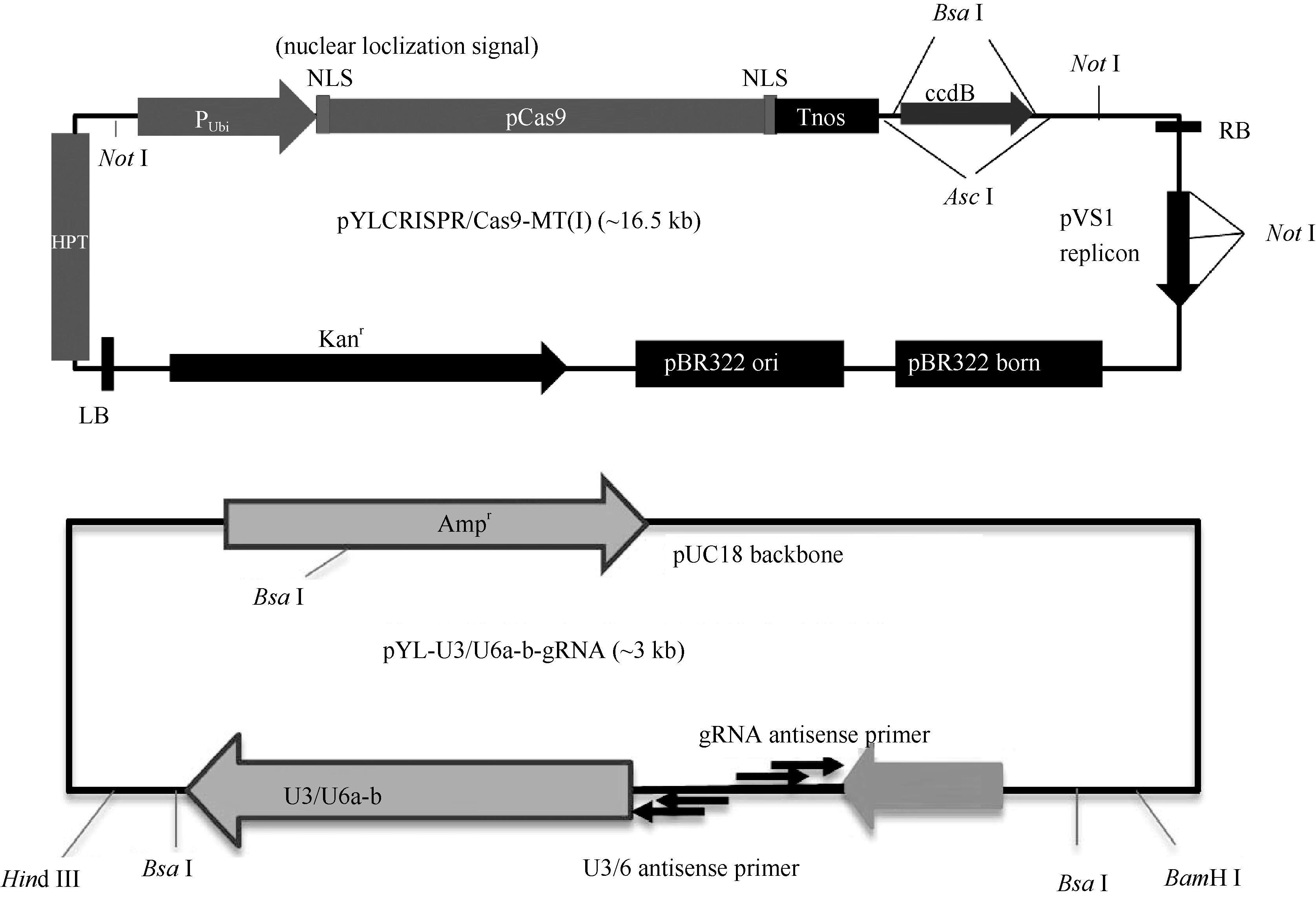
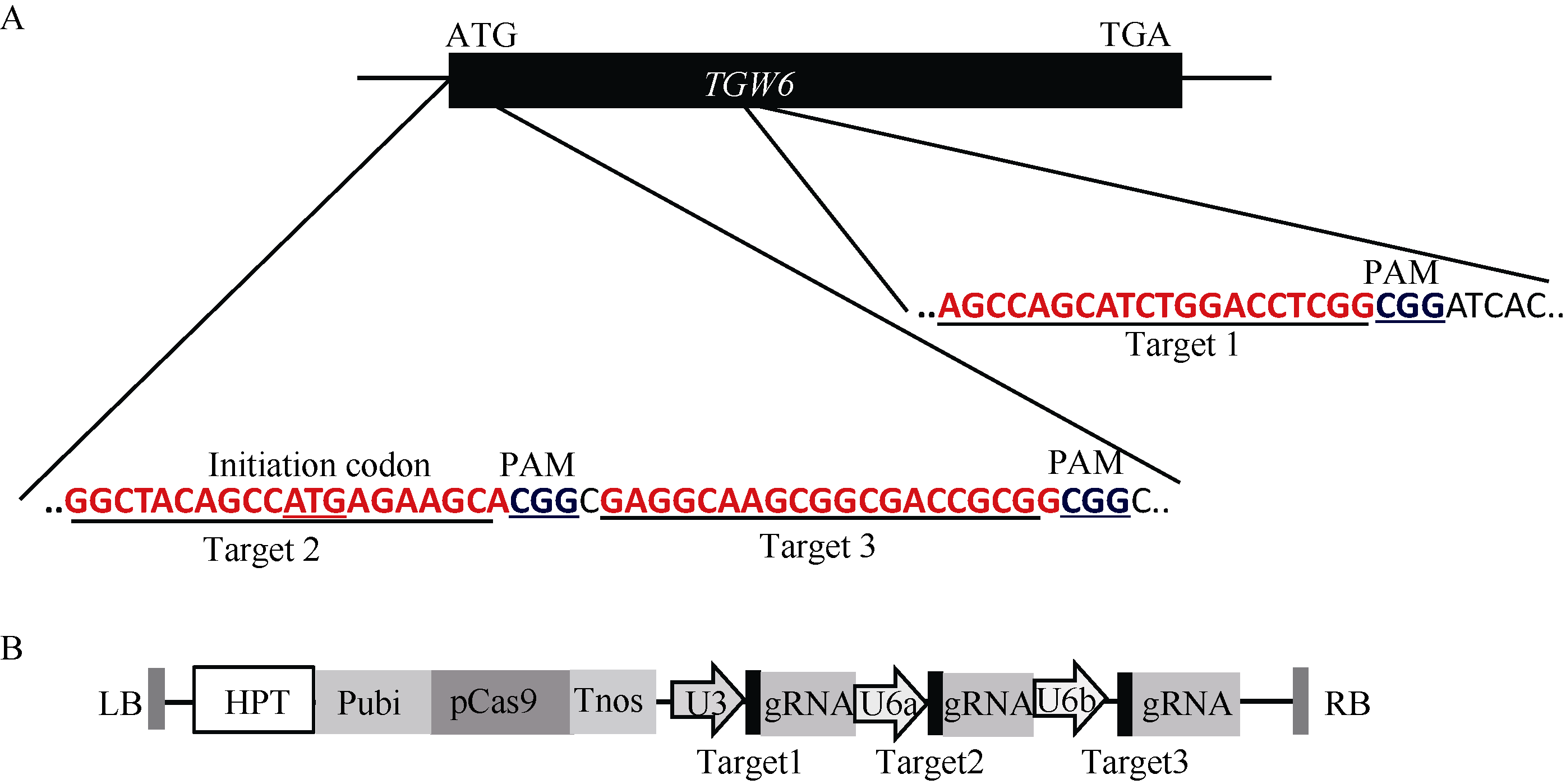
图2 gRNA靶位点及CRISPR/Cas9-gRNA的组装示意图 A: 3个靶点分别在TGW6基因内的位置; B: 3个靶点组装到pYLCRISPR/Cas9-MT(I)载体而成的T-DNA元件。Fig. 2 Target sites of the gRNA and cloning of gRNA cassette into the CRISPR/Cas9 vector A: positions of three targets in the TGW6 gene locus; B: T-DNA fragment assembled with the three targets and the pYLCRISPR/Cas9-MT(I) vector.
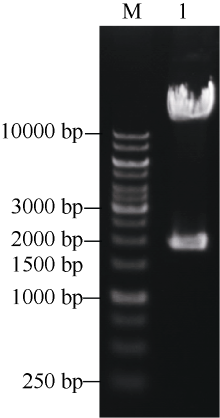
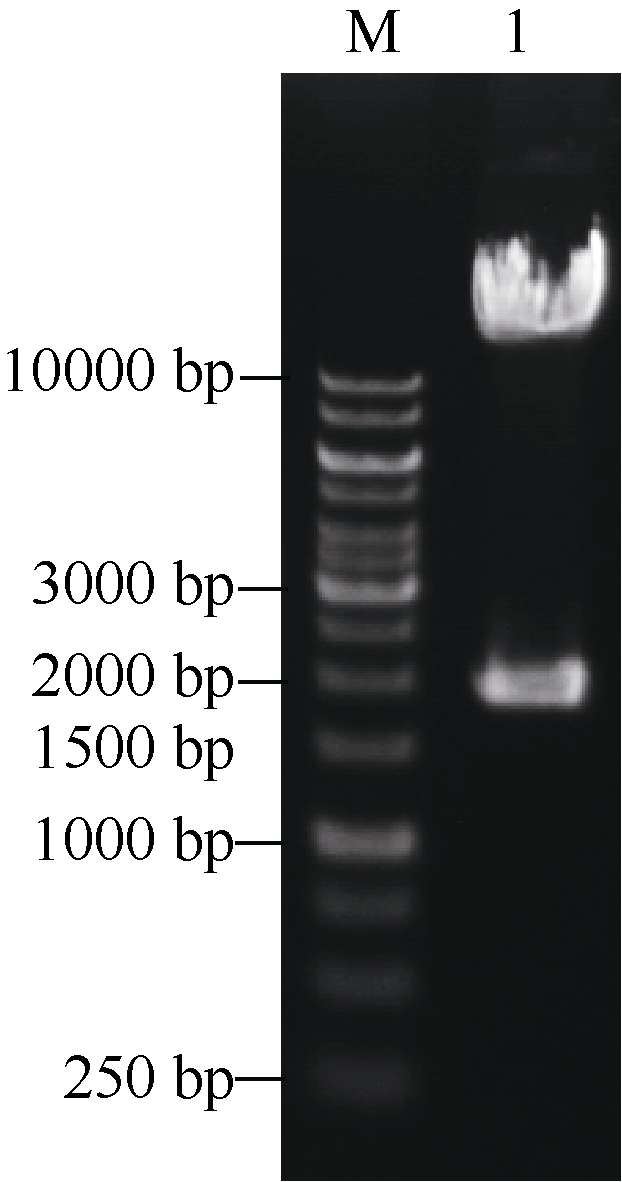
图3AscI酶切鉴定pYLCRISPR/Cas9-tgw6-gRNA载体Fig. 3 Identification of the pYLCRISPR/Cas9-tgw6-gRNA plasmid digested with Asc I M: 1 kb DNA ladder marker; 1: pYLCRISPR/Cas9-tgw6-gRNA.
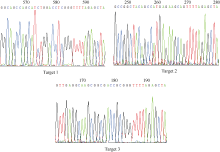
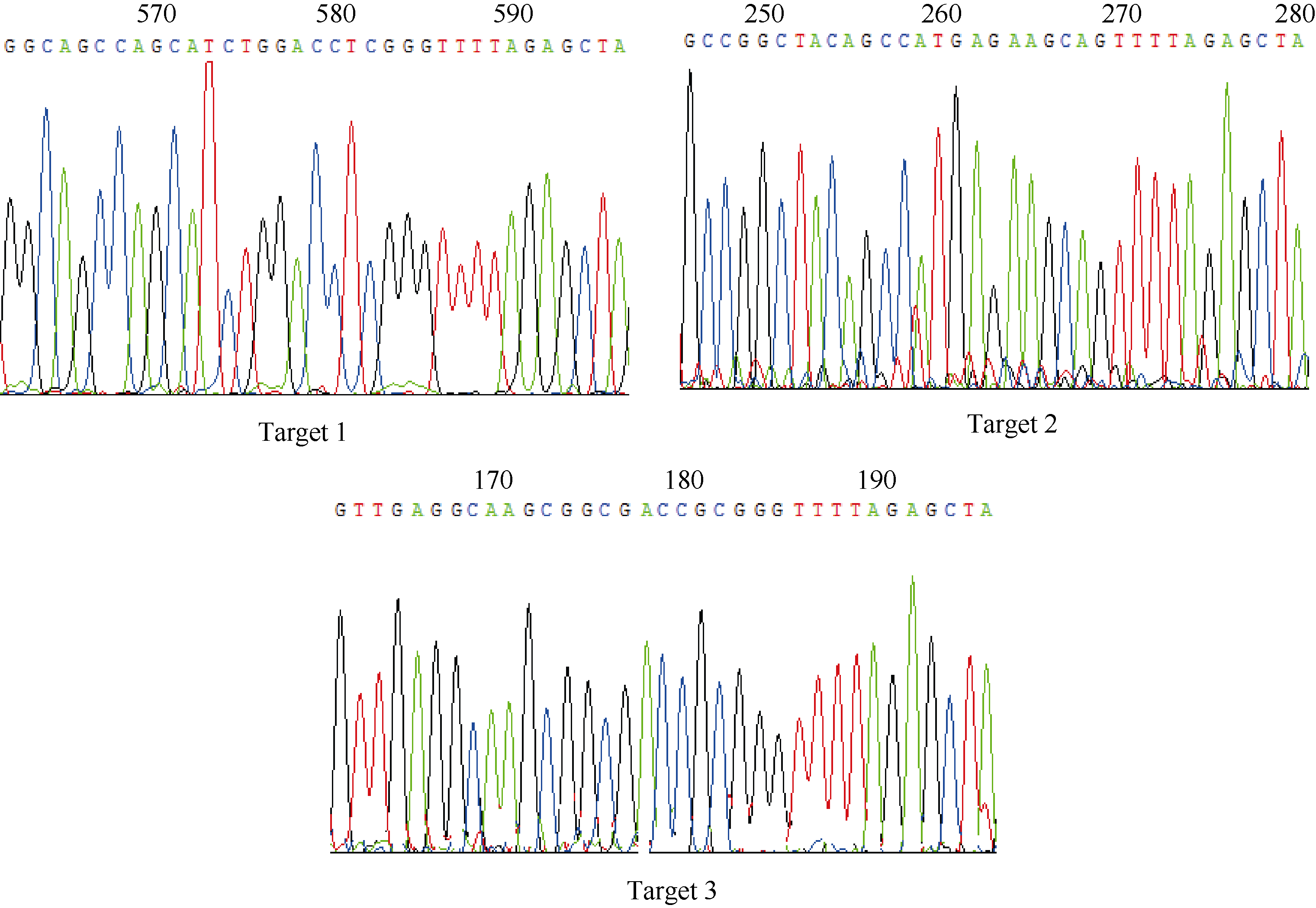
图5tgw6突变体的PCR检测及其与野生型序列比对分析 A: T0代(1~22)水稻tgw6编辑位点附近DNA片段的PCR检测结果, 野生型(WT)为H447, 水稻扩增长度为953 bp; B: 对应于A的PCR产物的测得序列与野生型(WT)的序列比对结果。“ Frequency” 指A电泳图对应的突变体中同一类型编辑位点突变个体出现的频率。Fig. 5 PCR identification and sequence alignment of tgw6 mutants compared to the WT line A: PCR identification results for the DNA fragments near the edited locus of thetgw6 T0 mutants (1-22) and WT line (H447); B: sequence alignment of tgw6 mutants compared to the WT line. “ Frequency” refers to the frequency of the same edited mutation among the corresponding mutants in the electrophoresis map of A.
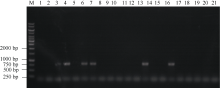
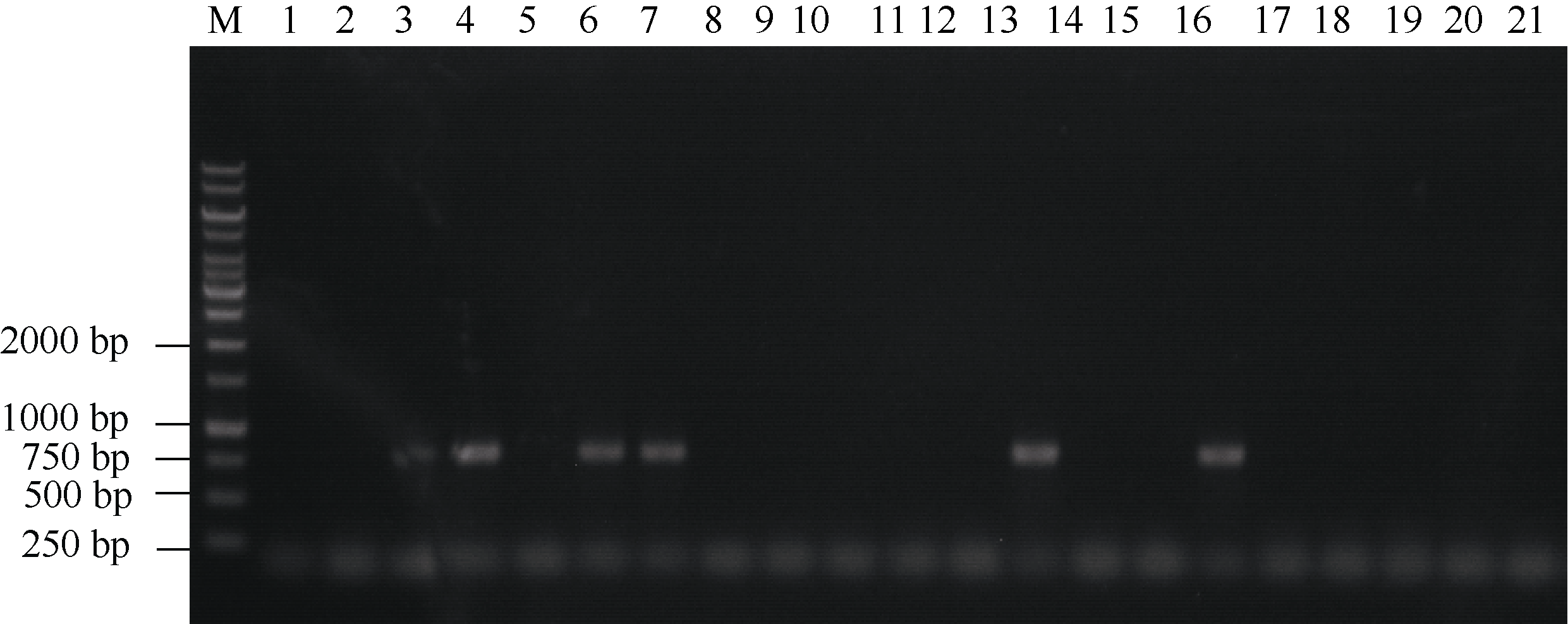
图6 部分tgw6突变体潮霉素基因的PCR检测 1: WT; 2~5: 缺失103 bp的突变体; 6~9: 缺失123 bp的突变体; 10~13: 缺失141 bp的突变体; 14~17: 缺失144 bp的突变体; 18~21: 缺失148 bp的突变体。Fig. 6 PCR identification for hph gene of parts of tgw6 mutants 1: WT; 2-5: the 103-bp deletion-mutants; 6-9: the 123-bp deletion-mutants; 10-13: the 141-bp deletion-mutants; 14-17: the 144-bp deletion-mutants; 18-21: the 148-bp deletion-mutants.
表2 Table 2 表2(Table 2)
表2 不同类型tgw6缺失突变体的千粒重测定结果 Table 2 Thousand grain weight of different types of homozygous deletion mutants of tgw6
突变类型 Mutant type
缺失碱基数 Base deletion (bp)
千粒重 Thousand-grain weight (g)
增加比例 Percentage increased (%)
野生型Wide type
0
22.0± 0.50 a
0
1
103
23.1± 0.57 b
5.0
2
123
23.4± 0.50 b
6.3
3
141
23.1± 0.47 b
5.0
4
144
23.7± 0.38 b
7.7
5
148
24.0± 0.35 b
8.2
Data listed in the table are mean± standard error; values followed by the same letter within the same column are not significantly different at the 0.05 probability level (Duncan’ s method). 表中所列数据为平均值± 标准误; 表中同列数据后字母相同者表示在0.05水平上差异不显著(Duncan’ s法)
表2 不同类型tgw6缺失突变体的千粒重测定结果 Table 2 Thousand grain weight of different types of homozygous deletion mutants of tgw6
You AQ, Lu XG, Jin HJ, RenX, LiuK, Yang GC, Yang HY, Zhu LL, He GC. Identification of quantitative trait loci across recombinant inbred lines and testcross populations for traits of agronomic importance in rice. Genetics, 2006, 172: 1287-1300[本文引用:1]
[2]
ShomuraA, IzawaT, EbanaK, EbitaniT, KanegaeH, KonishiS, YanoM. Deletion in a gene associated with grain size increased yields during rice domestication. Nat Genet, 2008, 40: 1023-1028[本文引用:1]
[3]
Weng JF, Gu SH, Wan XY, GaoH, GuoT, SuN, LeiC, ZhangX, Cheng ZJ, Guo XP, Wang JL, JiangL, Zhai HQ, Wan JM. Isolation and initial characterization of GW5, a major QTL associated with rice grain width and weight. Cell Res, 2008, 18: 1199-1209[本文引用:1]
[4]
IshimaruK, HirotsuN, MadokaY, MurakamiN, HaraN, OnoderaH, KashiwagiT, UjiieK, ShimizuB, OnishiA, MiyagawaH, KatohE. Loss of function of the IAA-glucose hydrolase gene TGW6 enhances rice grain weight and increases yield. Nat Genet, 2013, 45: 707-711[本文引用:2]
[5]
Fan CC, Xing YZ, Mao HL, Lu TT, HanB, Xu CG, Li XH, Zhang QF. GS3, a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein. Theor Appl Genet, 2006, 112: 1164-1171[本文引用:1]
[6]
Li YB, Fan CC, Xing YZ, Jiang YH, Luo LJ, SunL, ShaoD, Xu CJ, LiX, Xiao JH, He YQ, Zhang QF. Natural variation in GS5 plays an important role in regulating grain size and yield in rice. Nat Genet, 2011, 43: 1266-1269[本文引用:1]
[7]
Song XJ, HuangW, ShiM, Zhu MZ, Lin HX. A QTL for rice grain width and weight encodes a previously unknown RING-type E3 ubiquitin ligase. Nat Genet, 2007, 39: 623-630[本文引用:1]
[8]
Wang SK, WuK, YuanQ, LiuX, LiuZ, LinX, ZengR, ZhuH, DongG, QianQ, Zhang GQ, Fu XD. Control of grain size, shape and quality by OsSPL16 in rice. Nat Genet, 2012, 44: 950-954[本文引用:1]
[9]
Hu ZJ, He HH, Zhang SY, SunF, XinX, WangW, QianX, Yang JS, Luo XJ. A Kelch motif—containing serine/threonine protein phosphatase determines the large grain QTL trait in rice. J Integr Plant Biol, 2012, 54: 979-990[本文引用:2]
[10]
QiP, Lin YS, Song XJ, Shen JB, HuangW, Shan JX, Zhu MZ, JiangL, Gao JP, Lin HX. The novel quantitative trait locus GL3. 1 controls rice grain size and yield by regulating Cyclin-T1; 3. Cell Res, 2012, 22: 1666-1680[本文引用:1]
[11]
ZhangX, WangJ, HuangJ, LanH, WangC, YinC, WuY, TangH, QianQ, LiJ, ZhangH. Rare allele of OsPPKL1 associated with grain length causes extra-large grain and a significant yield increase in rice. Proc Natl Acad Sci USA, 2012, 109: 21534-21539[本文引用:1]
[12]
RathD, AmlingerL, RathA, LundgrenM. The CRISPR-Cas immune system: biology, mechanisms and applications. Biochimie, 2015, 117: 119-128[本文引用:1]
OsakabeY, OsakabeK. Genome editing with engineered nucleases in plants. Plant Cell Physiol, 2015, 56: 389-400[本文引用:1]
[15]
JiangW, BikardD, CoxD, ZhangF, Marraffini LA. RNA-guided editing of bacterial genomes using CRISPR-Cas systems. Nat Biotechnol, 2013, 31: 233-239[本文引用:1]
[16]
FengZ, ZhangB, DingW, LiuX, Yang DL, WeiP, CaoF, ZhuS, ZhangF, MaoY, Zhu JK. Efficient genome editing in plants using a CRISPR/Cas system. Cell Res, 2013, 23: 1229-1232[本文引用:1]
[17]
BortesiL, FischerR. The CRISPR/Cas9 system for plant genome editing and beyond. Biotechnol Adv, 2015, 33: 41-52[本文引用:1]
[18]
Ma XL, ZhangQ, ZhuQ, LiuW, ChenY, QiuR, WangB, YangZ, LiH, LinY, XieY, ShenR, ChenS, WangZ, ChenY, GuoJ, ChenL, ZhaoX, DongZ, Liu YG. A robust CRISPR/Cas9 system for convenient, high-efficiency multiplex genome editing in monocot and dicotplants. Mol Plant, 2015, 8: 1274-1284[本文引用:3]
[19]
Xu RF, LiH, Qin RY, LiJ, Qiu CH, Yang YC, MaH, LiL, Wei PC, Yang JB. Generation of inheritable and “transgene clean” targeted genome-modified rice in later generations using the CRISPR/Cas9 system. Sci Rep, 2015, 11491, doi: DOI:10.1038/srep11491[本文引用:3]
[20]
HieiY, OhtaS, KomariT, KumashiroT. Efficient transformation of rice (Oryza sativa L. ) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J, 1994, 6: 271-282[本文引用:1]
[21]
WangH, ChuZ, MaX, LiR, LiuY. A high through-Put protocol of plant genomic DNA preparation for PCR. Acta Agron Sin, 2013, 39: 1200-1205[本文引用:1]
[22]
BibikovaM, GolicM, Golic KG, CarrollD. Targeted chromosomal cleavage and mutagenesis in Drosophila using zinc-finger nucleases. Genetics, 2002, 161: 1169-1175[本文引用:1]
[23]
BibikovaM, BeumerK, Trautman JK, CarrollD. Enhancing gene targeting with designed zinc finger nucleases. Science, 2003, 300: 764[本文引用:1]
[24]
DreierB, Fuller RP, Segal DJ, Lund CV, BlancafortP, HuberA, KokschB, Barbas CF. Development of zinc finger domains for recognition of the 5′-CNN-3′ family DNA sequences and their use in the construction of artificial transcription factors. J Biol Chem, 2005, 280: 35588-35597[本文引用:1]
[25]
HockemeyerD, WangH, KianiS, Lai CS, GaoQ, Cassady JP, Cost GJ, ZhangL, SantiagoY, Miller JC, ZeitlerB, Cherone JM, MengX, Hinkley SJ, Rebar EJ, Gregory PD, Urnov FD, JaenischR. Genetic engineering of human pluripotent cells using TALE nucleases. Nat Biotechnol, 2011, 29: 731-734[本文引用:1]
[26]
TessonL, UsalC, MénoretS, LeungE, Niles BJ, RemyS, SantiagoY, Vincent AI, MengX, ZhangL, Gregory PD, AnegonI, Cost GJ. Knockout rats generated by embryo microinjection of TALENs. Nat Biotechnol, 2011, 29: 695-696[本文引用:1]
EndoM, MikamiM, TokiS. Multigene knockout utilizing off-target mutations of the CRISPR/Cas9 system in rice. Plant Cell Physiol, 2015, 56: 41-47[本文引用:1]
[29]
JiangW, ZhouH, BiH, FrommM, YangB, Weeks DP. Demonstration of CRISPR/Cas9/sgRNA-mediated targeted gene modification in Arabidopsis, tobacco, sorghum and rice. Nucl Acids Res, 2013, 41: e188. doi: DOI:10.1093/nar/gkt780[本文引用:1]
[30]
DiCarlo J E, Norville J E, Mali P, Rios X, Aach J, Church G M. Genome engineering in Saccharomyces cerevisiae using CRISPR-Cas systems. Nucl Acids Res, 2013, 41: 4336-4343[本文引用:1]
Gratz SJ, Cummings AM, Nguyen JN, Hamm DC, Donohue LK, Harrison MM, WildongerJ, O’Connor-Giles K M. Genome engineering of Drosophila with the CRISPR RNA guided Cas9 nuclease. Genetics, 2013, 194: 1029-1035[本文引用:1]
 , 郑才敏, 刘维, 罗文龙, 王慧, 陈志强
, 郑才敏, 刘维, 罗文龙, 王慧, 陈志强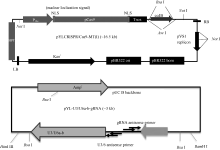



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}