关键词:水稻( Oryza sativaL.); 斑马叶突变体; 叶绿体; 基因精细定位 Morphological Characterization and Fine Mapping of Zebra Leaf Mutant zebra1349in Rice ( Oryza sativaL.) GUO Guo-Qiang1,2,3, SUN Xue-Wu2, SUN Ping-Yong2, YIN Jian-Ying3, HE Qiang2, YUAN Ding-Yang2,*, DENG Hua-Feng1,2,*, YUAN Long-Ping1,2,* 1College of Agronomy, Hunan Agricultural University, Changsha 410128, China
2State Key Laboratory of Hybrid Rice, Hunan Hybrid Rice Research Center, Changsha 410125, China
3Hengyang Agricultural Science Research Institute, Hengyang 421001, China
Fund:The study was supported by the grants from National High-tech R&D Program of China (863 Program)(2011AA10A101) and the Key Project Funded by the Hengyang Science and Technology Bureau (2011KZ15) AbstractA new zebra leaf mutant zebra1349was attained in a restorer line crossing population of [R128//(R318/R1025) F1] F6 in Hengyang Agricultural Science Research Institute of Hunan province. This mutant showed normal green leaves at seedlings stage, but a zebra leaf phenotype with green-yellow bands in penpendicular to leaf vein appeared at five days after transplanting, which was most obvious at sixth-leaf stage, and recovered normal green leaves around 30 days (ninth-leaf stage) after transplanting. Until the mature stage, the zebra1349 mutant showed insignificant difference with the wild type in major agronomic traits. The contents of total chlorophyll, chlorophyll a, chlorophyll b and carotenoid in yellow parts of the mutant leaf at sixth-leaf stage decreased by 55.86%, 61.02%, 39.34%, and 47.03%, respectively. Transmission Electron Microscopic (TEM) results indicated that the chloroplast of the mutant yellow leaf showed a serious thylakoid membrane degradation and decomposition, and the number of thylakoid grana lamella decreased significantly with larger gap and looser arrangement. Genetic analysis using F1and F2of the reciprocal crosses between zebra1349 and normal green rice varieties revealed that the zebra-leaf trait was controlled by one pair of recessive nuclear genes. With 1192 recessive plants in a F2population from the cross between zebra1349 mutant and normal green variety 02428, the ZEBRA1349gene was finely mapped between two InDel markers indel39 and indel44 on chromosome 12 with a genetic distance of 0.04 cM and 0.17 cM respectively, and the physical distance was 89 kb based on comparing with the reference genome of Japonicarice Nipponbare. These results provide a foundation for further map-based cloning of ZEBRA1349 and molecular marker-assisted breeding.
Keyword:Rice ( Oryza sativa L.); Zebra leaf mutant; Chloroplast; Gene fine mapping Show Figures Show Figures
图1 突变体zebra1349与其野生型亲本R1349在不同时期的表型 A: 苗期, 移栽前; B: 移栽后5 d, 斑马叶出现; C: 移栽后30 d, 斑马叶复绿; D: 成熟期; WT: 野生型; M: 突变体。Fig. 1 Phenotype of thezebra1349 mutant and its wild-type parent R1349 at the different stages A: seedling stage, before transplanting; B: at five days (d) after transplanting, zebra leaves appeared; C: at thirty days (d) after transplanting zebra leaves disappeared; D: mature stage. WT: wild type; M: mutant.
表1 Table 1 表1(Table 1)
表1 突变体zebra1349与野生型(WT)亲本主要农艺性状比较 Table 1 Comparison of major agronomic traits between the zebra1349 mutant and its wild-type (WT) parent
材料 Material
生育期 Whole growth period (d)
株高 Plant height (cm)
剑叶长 Flag leaf length (cm)
有效穗数 No. of effective panicles
穗长 Panicle length (cm)
每穗总粒数 No. of spikelets per panicle
结实率 Seed setting rate (%)
千粒重1000-grain weigh (g)
WT
127.50± 1.31
88.71± 2.60
19.16± 1.19
7.94± 0.61
18.60± 0.76
148.40± 5.52
92.04± 1.31
22.1± 0.12
zebra1349
128.30± 1.42
87.45± 2.29
18.27± 1.23
8.07± 0.70
18.30± 0.52
146.46± 3.29
92.87± 0.83
22.2± 0.12
|t|
1.242
1.091
1.560
0.420
0.977
0.906
1.606
1.768
The zebra1349mutant and its wild-type had no significant difference in those traits investigated (P > 0.05), t(18)0.05=2.10. zebra1349和野生型的所有性状均差异不显著(P > 0.05), t(18)0.05=2.10。
表1 突变体zebra1349与野生型(WT)亲本主要农艺性状比较 Table 1 Comparison of major agronomic traits between the zebra1349 mutant and its wild-type (WT) parent
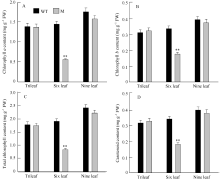
图2zebra1349和野生型(WT)不同发育时期叶片中色素含量分析 * * 表示野生型与突变体在0.01水平上差异显著。Fig. 2Pigment content in leaves of zebra1349 mutant and wild type (WT) at different growth stages * * Represent significant difference between zebra1349 mutant and the wild type at the 0.01 probability level.
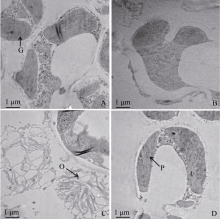
图3 突变体zebra1349和野生型叶肉细胞中叶绿体显微结构 A、B、C、D分别为野生型、zebra1349斑马叶绿区部位和黄区部位及复绿叶片的叶绿体结构。P: 原片层体; G: 基粒; O: 嗜锇粒。Fig. 3 Ultrastructures of chloroplasts in the mesophyll cell of thezebra1349 mutant and WT A, B, C, D represent chloroplasts of the wild type, the green parts, the yellow parts and green leaves of the zebra1349mutant, respectively. P: prolamellar body; G: granum; O: osmiophilic globule.
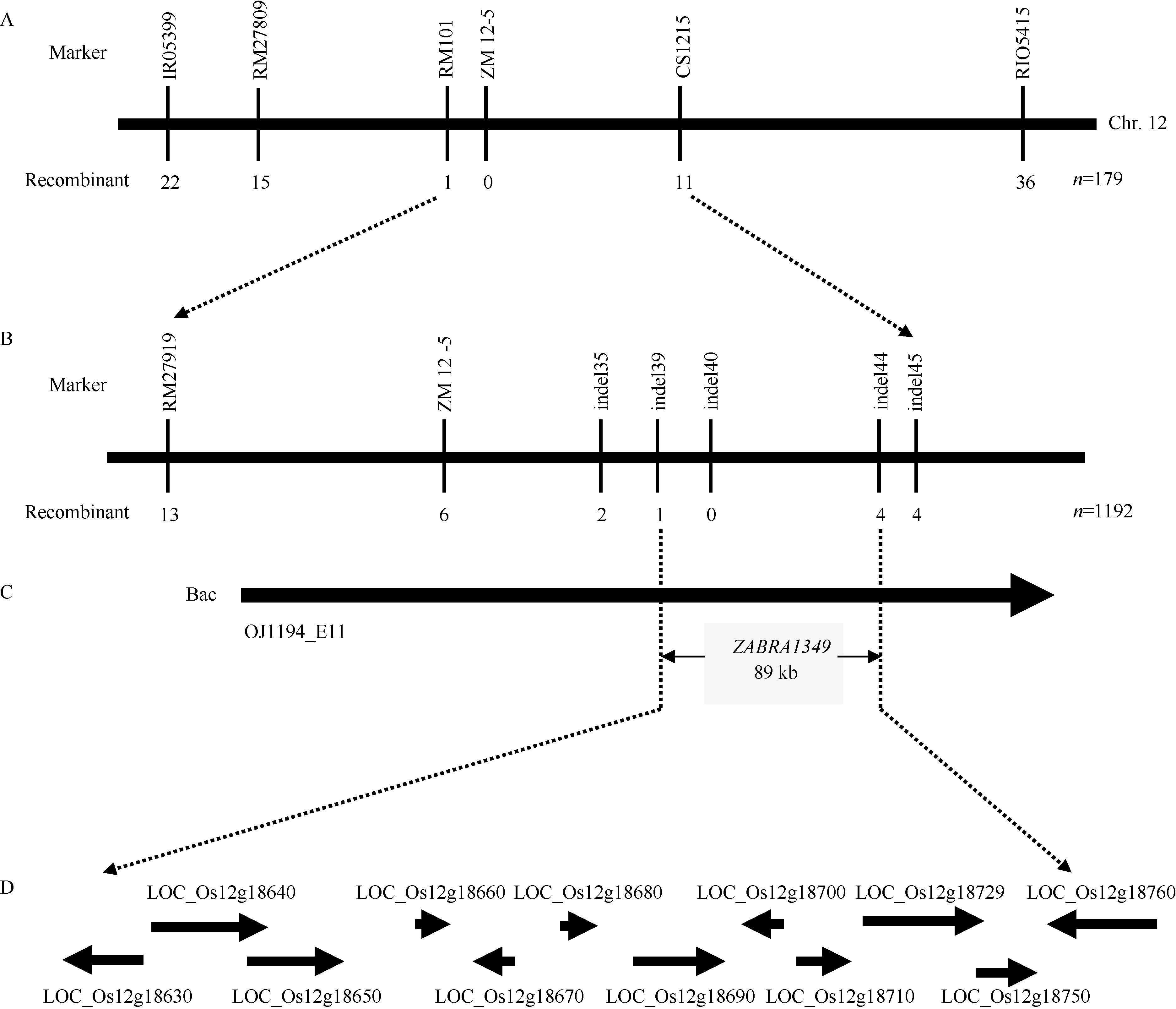
图4ZEBRA1349基因在第12染色体上的精细定位 A: ZEBRA1349被定位到第12染色体RM101与CS1215之间; B: ZEBRA1349被精细定位在InDel标记indel39与indel44之间; C: ZEBRA1349被定位在BAC克隆OJ1194_E11 89 kb范围内; D: 定位区间预测的基因。Fig. 4 Fine mapping ofZEBRA1349 on chromosome 12 A:ZEBRA1349 was positioned between RM101 and CS1215 on chromosome 12; B: ZEBRA1349was fine mapped between two InDel markers indel39 and indel44; C: ZEBRA1349was located in a BAC of OJ1194_E11 within 89 kb; D: putative genes in the target interval.
表4 Table 4 表4(Table 4)
表4 水稻第12染色体定位区间内基因及其推测功能 Table 4 Gene names and their putative functions in the target interval
基因名称 Gene name
推测功能 Putative function
LOC_Os12g18630
Expressed protein
LOC_Os12g18640
Pentatricopeptide, putative, expressed
LOC_Os12g18650
Regulator of chromosome condensation domain containing protein, expressed
LOC_Os12g18660
Expressed protein
LOC_Os12g18670
Expressed protein
LOC_Os12g18680
Retrotransposon protein, putative, unclassified
LOC_Os12g18690
Expressed protein
LOC_Os12g18700
Expressed protein
LOC_Os12g18710
Expressed protein
LOC_Os12g18729
Expressed protein
LOC_Os12g18750
Expressed protein
LOC_Os12g18760
Peptidase family C78 domain containing protein, expressed
表4 水稻第12染色体定位区间内基因及其推测功能 Table 4 Gene names and their putative functions in the target interval
Awan MA, Konzak CF, Rutger JN, Nilan RA. Mutagenic effects of sodium azide in rice. Crop Sci, 1980, 20: 663-668[本文引用:1]
[2]
黄晓群, 赵海新, 董春林, 孙业盈, 王平荣, 邓晓建. 水稻叶绿素合成缺陷突变体及其生物学研究进展. 西北植物学报, 2005, 25: 1685-1691Huang XQ, Zhao HX, Dong CL, Sun YY, Wang PR, Deng XJ. Chlorophyll-deficit rice mutants and their research advances in biology. Acta Bot Boreali-Occident Sin, 2005, 25: 1685-1691 (in Chinese with English abstract)[本文引用:1]
[3]
FambriniM, CastagnaA, Dalla VecchiaF, Degl’innocentiE, RanieriA, VernieriP, PardossiA, GuidiL, RascioN, PugliesiC. Characterization of a pigment-deficient mutant of sunflower (Helianthus annuus L. ) with abnormal chloroplast biogenesis, reduced PSII activity and low endogenous level of abscisic acid. Plant Sci, 2004, 167: 79-89[本文引用:1]
[4]
Parks BM, Quail PH. Phytochrome-deficient hy1 and hy2 long hypocotyl mutants of Arabidopsis are defective in phytochrome chromophore biologysynthesis. Plant Cell, 1991, 3: 1177-1186[本文引用:1]
[5]
Agrawal GK, YamazakiM, KobayashiM, HirochikaR, MiyaoA, HirochikaH. Screening of the rice viviparous mutants generated by endogenous retrotransposon tos17 insertion. Tagging of a zeaxanthin epoxidase gene and a novel OsTATC gene. Plant Physiol, 2001, 125: 1248-1257[本文引用:1]
[6]
Stern DB, Hanson MR, BarkanA. Genetics and genomics of chloroplast biogenesis: maize as a model system. Trends Plant Sci, 2004, 9: 293-301[本文引用:1]
[7]
沈圣泉, 舒庆尧, 吴殿星, 陈善福, 夏英武. 白化转绿型水稻三系不育系白丰A的选育. 杂交水稻, 2005, 20(5): 10-11Shen SQ, Shu QY, Wu DX, Chen SF, Xia YW. Breeding of new rice CMS line Baifeng A with a green-revertible albino leaf color marker. Hybrid Rice, 2005, 20(5): 10-11 (in Chinese with English abstract)[本文引用:1]
[8]
邓晓娟, 张海清, 王悦, 舒志芬, 王国槐, 王国梁. 水稻叶色突变基因研究进展. 杂交水稻, 2012, 27(5): 9-14Deng XJ, Zhang HQ, WangY, Shu ZF, Wang GH, Wang GL. Research advances on rice leaf-color mutant genes. Hybrid Rice, 2012, 27(5): 9-14 (in Chinese with English abstract)[本文引用:2]
[9]
谭炎宁, 孙学武, 袁定阳, 孙志忠, 余东, 何强, 段美娟, 邓华凤, 袁隆平. 水稻单叶独立转绿型黄化突变体grc2的鉴定与基因精细定位. 作物学报, 2015, 41: 831-837Tan YN, Sun XW, Yuan DY, Sun ZZ, YuD, HeQ, Duan MJ, Deng HF, Yuan LP. Identification and fine mapping of green-revertible chlorina gene grc2 in rice (Oryza sativa L. ). Acta Agron Sin, 2015, 41: 831-837 (in Chinese with English abstract)[本文引用:1]
[10]
钱前, 朱旭东, 曾大力, 张小惠, 严学强, 熊振民. 细胞质基因控制的新特异材料白绿苗的研究. 作物品种资源, 1996, (4): 11-12QianQ, Zhu XD, Zeng DL, Zhang XH, Yan XQ, Xiong ZM. The study on a new special material, white-green rice which controlled by plasma gene. J Crop Resour, 1996, (4): 11-12 (in Chinese)[本文引用:1]
[11]
李贤勇, 王楚桃, 李顺武, 何永歆, 陈世全. 一个水稻高叶绿素含量基因的发现. 西南农业学报, 2002, 15(4): 122-123Li XY, Wang CT, Li SW, He YY, Chen SQ. The discovery of a high chlorophyll content gene in rice. Southwest China J Agric Sci, 2002, 15(4): 122-123 (in Chinese)[本文引用:1]
[12]
Jung KH, HurJ, Ryu CH, ChoiY, Chung YY, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:1]
[13]
Zhang HT, Li JJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC. Rice chlorina-1 and chlorina-9 encode ChlD and ChlI subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1]
[14]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
[15]
LeeS, Kim JH, Yoo ES, Lee CH, HirohikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1]
[16]
SugimotoH, KusumiK, TozawaY, YazakiJ, KishimotoN, KikuchiS, IbaK. The virescent-2 mutation inhibits translation of plastid transcripts for the plastid genetic system at an early stage of chloroplast differentiation. Plant Cell Physiol, 2004, 45: 985-996[本文引用:1]
[17]
Yoo SC, Cho SH, SugimotoH, LiJ, KusumiK, Koh HJ, KohI, Paek NC. Rice virescent3 and stripe1 encoding the large and small subunits of ribonucleotide reductase are required for chloroplast biogenesis during early leaf development. Plant Physiol, 2009, 150: 388-401[本文引用:1]
[18]
Gothand am KM, Kim ES, Cho HJ, Chung YY. OsPPR1, a pentatricopeptide repeat protein of rice is essential for the chloroplast biogenesis. Plant Mol Biol, 2005, 58: 421-433[本文引用:1]
[19]
Park SY, Yu JW, Park JS, LiJ, Yoo SC, Lee NY, Lee SK, Jeong SW, Seo HS, Koh HJ, Jeon JS, Park YI, Paek NC. The senescence-induced stay green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1]
[20]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1]
[21]
YutakaS, RyouheiM, SusumuK, MinoruN, AyumiT, MakotoK. Two short-chain dehydrogenase/reductases, NON-YELLOW COLORING 1 and NYC1-LIKE, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J, 2009, 57: 120-131[本文引用:1]
[22]
Wang PR, Gao JX, Wan CM, Zhang FT, Xu ZJ, Huang XQ, Sun XQ, Deng XJ. Divinyl chlorophyll(ide) a can be converted to monovinyl chlorophyll(ide) a by a divinyl reductase in rice. Plant Physiol, 2010, 153: 994-1003[本文引用:]
[23]
Lichtenthaler HK. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1]
[24]
Murray MG, Thompson WF. Rapid isolation of high molecular weight plant DNA. Nucl Acids Res, 1980, 8: 4321-4326[本文引用:1]
[25]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1]
[26]
IwataN, OmuraT, SatoH. Linkage studies in rice (Oryza sativa L. ) on some mutants for physiological leaf spots. Fac Agric Kushu Univ, 1978, 22: 243-251[本文引用:1]
[27]
Wang QS, S C, Ling Y H, Zhao F M, Yang Z L, Li Y F, He G H. Genetic analysis and molecular mapping of a novel gene for zebra mutation in rice (Oryza sativa L. ). J Genet Genomics, 2009, 36: 679-684[本文引用:1]
[28]
李燕群, 钟萍, 高志艳, 朱柏羊, 陈丹, 孙昌辉, 王平荣, 邓晓建. 水稻斑马叶突变体zebra524的表型鉴定及候选基因分析. 中国农业科学, 2014, 47: 2907-2915Li YQ, ZhongP, Gao ZY, Zhu BY, ChenD, Sun CH, Wang PR, Deng XJ. Morphological characterization and cand idate gene analysis of zebra leaf mutant zebra524 in rice. Sci Agric Sin, 2014, 47: 2907-2915[本文引用:1]
[29]
Li JJ, Pand eyaD, NathK, Zulfugarov IS, Yoo SC, Zhang HT, Yoo JH, Cho SH, Koh HJon, Kim DS, Seo HS, Kang BC, Lee CH, Paek NC. ZEBRA-NECROSIS, a thylakoid-bound protein, is critical for the photoprotection of developing chloroplasts during early leaf development. Plant J, 2010, 62: 713-725[本文引用:1]
[30]
Chai CL, FangJ, LiuY, Tong HN, Gong YQ, Wang YQ, LiuM, Wang YP, QianQ, Cheng ZK, Chu CC. ZEBRA2, encoding a carotenoid isomerase, is involved in photo protection in rice. Plant Mol Biol, 2011, 75: 211-221[本文引用:1]
[31]
Mao DH, Yu HH, Liu TM, Yang GY, Xing YZ. Two complementary recessive genes in duplicated segments control etiolation in rice. Theor Appl Genet, 2011, 122: 373-383[本文引用:1]
[32]
Dong YJ, Lin DZ, MeiJ, Su QQ, Zhang JH, Ye SH, Zhang XM. Genetic analysis and molecular mapping of a thermo-sensitive chlorosis mutant in rice. Mol Plant Breed, 2013, 11: 1-7[本文引用:1]
[33]
Shi JQ, Wang YQ, GuoS, MaL, Wang ZW, Zhu XY, Sang XC, Ling YH, WangN, Zhao FM, He GH. Molecular mapping and cand idate gene analysis of a yellow-green leaf 6 (ygl6). Crop Sci, 2014, 55: 669-680[本文引用:1]
[34]
KusumiK, ChonoY, ShimadaH, GotohE, TsuyamaM, IbaK. Chloroplast biogenesis during the early stage of leaf development in rice. Plant Biotechnol, 2010, 27: 85-90[本文引用:1]
[35]
LurinC, AndresC, AubourgS, BellaouiM, BittonF, BruyereC, CabocheM, DebastC, GualbertoJ, HoffmannB, LecharnyA, Ret ML, Martin-Magniette M L, Mireau H, Peeters N, Renou J P, Szurek Boris, Taconnat L, Small I. Genome-wide analysis of arabidopsis pentatricopeptide repeat proteins reveals their essential role in organelle biogenesis. Plant Cell, 2004, 16: 2089-2103[本文引用:2]
[36]
SuN, Hu ML, Wu DX, Wu FQ, Fei GL, LanY, Chen XL, Shu XL, ZhangX, Guo XP, Cheng ZJ, Lei CL, Qi CK, JiangL, Wang HY, Wan JM. Disruption of a rice pentatricopeptide repeat protein causes a seedling-specific albino phenotype and its utilization to enhance seed purity in hybrid rice production. Plant Physiol, 2012, 159: 227-238[本文引用:1]
[37]
舒庆尧, 夏英武, 左晓旭, 刘贵付. 二系杂交水稻制繁种中利用标记辅助去杂技术. 浙江农业大学学报, 1996, 22(1): 56-60Shu QY, Xia YW, Zuo XX, Liu GF. Maker-assisted elimination of contamination on two-line hybrid rice seed production and multiplication. J Zhejiang Agric Univ, 1996, 22(1): 56-60 (in Chinese)[本文引用:1]
 , 邓华凤
, 邓华凤





{kind=link}
{kind=link}
{kind=link}
{kind=link}