关键词:油菜; 分枝角度; 理想株型; 主基因+多基因; 遗传效应 Genetic Effects of Branch Angle Using Mixture Model of Major Gene Plus Polygene in Brassica napus L. WANG Wen-Xiang, HU Qiong, MEI De-Sheng, LI Yun-Chang, ZHOU Ri-Jin, WANG Hui, CHENG Hong- Tao, FU Li, LIU Jia* Oil Crops Research Institute of Chinese Academy of Agricultural Sciences / Key Laboratory of Biology and Genetic Improvement of Oil Crops of the Ministry of Agriculture, Wuhan 430062, China Fund:This study was support by the Science and Technology Innovation Project of Chinese Academy of Agricultural Sciences (Group No.118), the Earmarked Fund for China Agriculture Research System (CARS-13), the Natural Science Foundation of China (31471535), the Natural Science Foundation of Hubei Province (2014CFB156) and the Hubei Agricultural Science and Technology Innovation Center AbstractBranch angle is one of important plant architectural traits in rapeseed ( Brassica napus), which is the basic requirement for high-density cultivation and mechanical harvesting. To reveal the genetic mechanism of branch angle in rapeseed, we produced six generations (P1, P2, F1, F2, BCP1, and BCP2) from a cross between loose-type 6098B and compact-type Purler to analyze genetic effects of branch angle in rapeseed by the mixed major gene plus polygene inheritance model. The branch angle showed a continuous normal distribution, where both the basal branch angle and the top branch angle were dominated by a pair of major gene with additive-dominant effects plus polygenes with additive-dominance-epistasis effects (D-0 model). The additive effect and dominant effect of major gene of the top branch angle were 4.939 and -4.156, the heritability in BCP1, BCP2, and F2 was 34.08%, 1.40%, and 14.99%, respectively for the major gene, and 24.43%, 61.72%, and 63.98%, respectively for the polygenes. The additive effect and dominant effect of major gene of the basal branch angle were 2.217 and -1.941, the heritability in BCP1, BCP2, and F2 was 7.86%, 1.24%, and 4.84%, respectively for the major gene, and 66.46%, 58.49%, and 73.96%, respectively for the polygenes. Results of this study found that the branch angle traits of 6098B×Purler is controlled by polygenes, but there is a major gene among them. The research may be useful for further understanding and genetic improvement of plant architecture in rapeseed.
Keyword: Brassica napus L.; Branch angle; Ideal plant architecture; Major gene plus polygene; Genetic effects Show Figures Show Figures
图1 6098B× Purler杂交组合顶枝角的频次分布 A: BCP1分离群体的频次分布; B: BCP2分离群体的频次分布; C: F2分离群体的频次分布。Fig. 1 Frequency distribution of top branch angle from 6098B× Purler A: Frequency distribution in the BCP1populations; B: Frequency distribution in the BCP2 populations; C: Frequency distribution in the F2 populations.
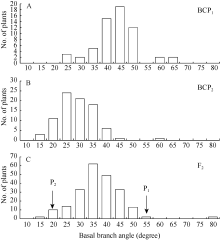
图2 6098B× Purler杂交组合基枝角的频次分布 A: BCP1分离群体的频次分布; B: BCP2分离群体的频次分布; C: F2分离群体的频次分布。Fig. 2 Frequency distribution of basal branch angle from 6098B× Purler A: Frequency distribution in the BCP1populations; B: Frequency distribution in the BCP2 populations; C: Frequency distribution in the F2 populations.
表2 6098B× Purler组合后代各遗传模型的AIC值 Table 2 AIC values of the genetic models from 6098B× Purler
模型代码 Model code
模型含义 Implication of model
AIC值 AIC value
极大似然函数值log_Max_likelihood_value
顶枝角 Top branch angle
基枝角 Basal branch angle
顶枝角 Top branch angle
基枝角 Basal branch angle
A-1
1MG-AD
2891.41
2801.24
-1441.70
-1396.62
A-2
1MG-A
2896.16
2806.81
-1445.08
-1400.40
A-3
1MG-EAD
2919.70
2796.71
-1456.85
-1395.35
A-4
1MG-AEND
2912.20
2858.57
-1453.10
-1426.29
B-1
2MG-ADI
2836.36
2717.54
-1408.18
-1348.77
B-2
2MG-AD
2879.57
2771.67
-1433.78
-1379.83
B-3
2MG-A
2885.88
2738.07
-1438.94
-1365.04
B-4
2MG-EA
2866.24
2737.06
-1430.12
-1365.53
B-5
2MG-AED
2973.21
2797.38
-1482.61
-1394.69
B-6
2MG-EEAD
2971.21
2795.38
-1482.61
-1394.69
C-0
PG-ADI
2846.25
2735.20
-1413.13
-1357.60
C-1
PG-AD
2860.88
2711.22
-1423.44
-1348.61
D-0
MX1-AD-ADI
2832.34
2704.68
-1404.17
-1340.34
D-1
MX1-AD-AD
2863.60
2710.13
-1422.80
-1346.06
D-2
MX1-A-AD
2860.09
2708.13
-1422.05
-1346.07
D-3
MX1-EAD-AD
2862.28
2708.21
-1423.14
-1346.11
D-4
MX1-AEND-AD
2851.54
2707.49
-1417.77
-1345.74
E-0
MX2-ADI-ADI
2840.74
2713.49
-1402.37
-1338.75
E-1
MX2-ADI-AD
2835.83
2706.85
-1402.92
-1338.43
E-2
MX2-AD-AD
2868.25
2712.78
-1423.12
-1345.39
E-3
MX2-A-AD
2847.70
2706.95
-1414.85
-1344.48
E-4
MX2-EA-AD
2862.21
2708.11
-1423.10
-1346.05
E-5
MX2-AED-AD
2864.28
2710.21
-1423.14
-1346.11
E-6
MX2-EEAD-AD
2862.28
2708.22
-1423.14
-1346.11
MG: major gene model; MX: mixed major gene and polygene model; PG: polygene model; A: additive effect; D: dominance effect; I: interaction; N: negative; E: equal; e.g. Model E-1 = MX2-ADI-AD, means mixed model with two major genes of additive-dominance- epistasis effects plus additive-dominance polygene. Minimum AIC value in each cross is underlined, showing the good-fitting model. MG: 主基因模型; MX: 主基因+多基因混合模型; PG: 多基因遗传模型; A: 加性效应; D: 显性效应; I: 互作; N: 负向; E: 相等; 例如E-1模型MX2-ADI-AD, 表示2对加性-显性-上位性主基因+加性-显性多基因混合遗传模型; 下画线表示AIC值最小, 用于最适遗传模型检验。
表2 6098B× Purler组合后代各遗传模型的AIC值 Table 2 AIC values of the genetic models from 6098B× Purler
表3 Table 3 表3(Table 3)
表3 6098B× Purler群体适合性检验 Table 3 Fitness tests of selected models in 6098B× Purler
性状Trait
模型Model
世代Generation
U12
U22
U32
nW2
Dn
顶枝角 Top branch angle
D-0
P1
0.016(0.898)
0.002(0.966)
0.106(0.745)
0.053(0.857)
0.113(1.000)
F1
0.087(0.768)
0.189(0.664)
0.353(0.553)
0.074(0.736)
0.046(1.000)
P2
0.001(0.977)
0.008(0.928)
0.221(0.639)
0.032(0.970)
0.059(1.000)
BCP1
0.002(0.967)
0.015(0.902)
0.108(0.743)
0.035(0.995)
0.009(1.000)
BCP2
0.250(0.617)
0.937(0.333)
3.750(0.053)
0.221(0.234)
0.012(1.000)
F2
0.095(0.758)
0.527(0.468)
2.924(0.087)
0.176(0.320)
0.005(1.000)
E-1
P1
1.415(0.234)
1.607(0.205)
0.215(0.643)
0.206(0.258)
0.155(0.997)
F1
0.003(0.957)
0.014(0.906)
0.464(0.496)
0.060(0.819)
0.047(1.000)
P2
0.027(0.871)
0.002(0.969)
0.228(0.633)
0.034(0.960)
0.063(1.000)
BCP1
0.003(0.959)
0.045(0.832)
0.420(0.517)
0.045(0.907)
0.026(1.000)
BCP2
0.053(0.818)
0.495(0.482)
3.692(0.055)
0.180(0.310)
0.012(1.000)
F2
0.036(0.849)
0.282(0.595)
1.929(0.165)
0.135(0.442)
0.005(1.000)
基枝角 Basal branch angle
D-0
P1
0.058(0.810)
0.030(0.862)
0.056(0.813)
0.137(0.436)
0.277(0.849)
F1
0.142(0.707)
0.021(0.885)
0.771(0.380)
0.101(0.593)
0.067(1.000)
P2
0.063(0.802)
0.037(0.847)
0.041(0.840)
0.062(0.805)
0.041(1.000)
BCP1
0.010(0.922)
0.073(0.788)
2.119(0.145)
0.142(0.416)
0.011(1.000)
BCP2
0.002(0.969)
0.002(0.963)
0.116(0.733)
0.038(0.944)
0.007(1.000)
F2
0.000(0.998)
0.247(0.619)
3.979(0.046)*
0.161(0.360)
0.005(1.000)
E-1
P1
0.475(0.491)
0.533(0.465)
0.063(0.801)
0.183(0.304)
0.207(0.982)
F1
0.301(0.583)
0.108(0.743)
0.662(0.416)
0.120(0.501)
0.062(1.000)
P2
0.014(0.908)
0.006(0.939)
0.021(0.885)
0.053(0.858)
0.038(1.000)
BCP1
0.024(0.877)
0.337(0.562)
2.960(0.085)
0.180(0.311)
0.017(1.000)
BCP2
0.035(0.851)
0.035(0.852)
0.001(0.982)
0.036(0.953)
0.007(1.000)
F2
0.171(0.680)
0.000(0.995)
2.478(0.115)
0.143(0.413)
0.005(1.000)
The numbers in brackets are the distribution values in theory. * Significant difference at the 0.05 probability level. U12、U22、U32栏中括号内数字为理论分布值。* 表示差异达到显著水平(P< 0.05)。
表3 6098B× Purler群体适合性检验 Table 3 Fitness tests of selected models in 6098B× Purler
王汉中, 殷艳. 我国油料产业形势分析与发展对策建议. 中国油料作物学报, 2014, 36: 414-421Wang HZ, YinY. Analysis and strategy for oil crop industry in China. Chin J Oil Crop Sci, 2014, 36: 414-421 (in Chinese with English abstract)[本文引用:1]
[2]
唐敏强, 程晓晖, 童超波, 刘越英, 赵传纪, 董彩华, 于景印, 马小艮, 黄军艳, 刘胜毅. 甘蓝型油菜株高性状的全基因组关联分析. 作物学报, 2015, 41: 1121-1126Tang MQ, Cheng XH, Tong CB, Liu YY, Zhao CJ, Dong CH, Yu JY, Ma XG, Huang JY, Liu SY. Genome-wide association analysis of plant height in rapeseed (Brassica napus). Acta Agron Sin, 2015, 41: 1121-1126 (in Chinese with English abstract)[本文引用:1]
[3]
王嘉, 荆凌云, 荐红举, 曲存民, 谌利, 李加纳, 刘列钊. 甘蓝型油菜株高、第一分枝高和分枝数的QTL检测及候选基因筛选. 作物学报, 2015, 41: 1027-1038WangJ, Jing LY, Jian HJ, Qu CM, ChenL, Li JN, Liu LZ. Quantitative trait loci mapping for plant height, the first branch height, and branch number and possible cand idate genes screening in Brassica napus L. Acta Agron Sin, 2015, 41: 1027-1038 (in Chinese with English abstract)[本文引用:1]
[4]
张凤启, 刘越英, 程晓辉, 童超波, 董彩华, 唐敏强, 黄军艳, 刘胜毅. 利用高密度SNP标记定位甘蓝型油菜株高QTL. 中国油料作物学报, 2014, 36: 695-700Zhang FQ, Liu YY, Cheng XH, Tong CB, Dong CH, Tang MQ, Huang JY, Liu SY. QTL mapping of plant heigh using high desity SNP markers in Brassica napus. Chin J Oil Crop Sci, 2014, 36: 695-700 (in Chinese with English abstract)[本文引用:1]
[5]
谢田田, 陈玉波, 黄吉祥, 张尧锋, 徐爱遐, 陈飞, 倪西源, 赵坚义. 甘蓝型油菜不同发育时期株高QTL的动态分析. 作物学报, 2012, 38: 1802-1809Xie TT, Chen YB, Huang JX, Zhang YF, Xu AX, ChenF, Ni XY, Zhao JY. Dynamic analysis of QTL for plant height of rapeseed at different developmental stages. Acta Agron Sin, 2012, 38: 1802-1809 (in Chinese with English abstract)[本文引用:1]
[6]
Xu JS, SongX, ChengY, Zou XL, ZengL, QiaoX, Lu GY, Fu GP, QuZ, Zhang XK. Identification of QTLs for branch number in oilseed rape (Brassica napus L. ). J Genet Genom, 2014, 41: 557-559[本文引用:1]
[7]
LiF, Chen BY, XuK, Gao GZ, Yan GX, Qiao JW, LiJ, LiH, Li LX, Zhang TY, TakeshiNishio, Wu XM. A genome-wide association study of plant height and primary branch number in rapeseed (Brassica napus). Plant Sci, 2015, 242: 169-177[本文引用:1]
[8]
王俊生, 李少钦, 张耀文. 不同栽培密度对紧凑型油菜产量和主要性状的影响. 耕作与栽培, 2006, (3): 25-26Wang JS, Li SQ, Zhang YW. Effects of different planting density on yield and main characters of compact in rapeseed. Gengzuo yu Zaipei, 2006, (3): 25-26 (in Chinese)[本文引用:1]
[9]
张倩. 甘蓝型油菜主要株型性状的遗传分析和QTL初步定位. 西南大学硕士学位论文, 重庆, 2013. pp 3-7ZhangQ. Genetic Effects Analysis and QTL Mapping of Major Plant-Type Traits in Brassica napus L. MS Thesis of Southwest University, Chongqing, China, 2013. pp 3-7 (in Chinese with English abstract)[本文引用:2]
[10]
张文学, 李殿荣, 田建华. 甘蓝型油菜紧凑型和松散型杂种一代分枝性状的遗传效应. 中国农学通报, 2000, 16(5): 17-19Zhang WX, Li DR, Tian JH. Genetic effect of hybrid F1 branching character of CCT and LCT in Brassica napus. Chin Agric Sci Bull, 2000, 16(5): 17-19 (in Chinese with English abstract)[本文引用:3]
[11]
王俊生, 田建华, 张继澍, 李殿荣, 袁志发. 紧凑型油菜株型性状的遗传及其与主要产量性状的相关性研究. 西北农林科技大学学报(自然科学版), 2005, 33: 7-12Wang JS, Tian JH, Zhang JS, Li DR, Yuan ZF. Study on the heredity of plant-type traits in compact rapeseed lines and the genetic correlation with yield traits. J Northwest Sci-Tech Univ Agric For (Nat Sci Edn), 2005, 33: 7-12 (in Chinese with English abstract)[本文引用:4]
[12]
唐佳. 甘蓝型油菜株型相关性状的杂种优势及配合力分析. 四川农业大学硕士学位论文, 四川成都, 2012. pp 3-7TangJ. Analyses on Heterosis and Combining Ability of the Traits Related to Plant Types in Brassica napus L. MS Thesis of Sichuan Agricultural University, Sichuan, China, 2014. pp 3-7 (in Chinese with English abstract)[本文引用:1]
[13]
漆丽萍. 甘蓝型油菜株型与角果相关性状的QTL分析. 华中农业大学博士学位论文, 湖北武汉, 2014. pp 11-12Qi LP. QTL Analysis for Traits Associated with Plant Architecture and Silique in Brassica napus L. PhD Dissertation of Huazhong Agricultural University, Wuhan, China, 2014. pp 11-12 (in Chinese with English abstract)[本文引用:1]
[14]
LiuJ, WangW, MeiD, WangH, FuL, LiuD, LiY, HuQ. Characterising variation of branch angle and genome-wide association mapping in rapeseed (Brassica napus L. ). Front Plant Sci, 2016, 7: 21[本文引用:1]
[15]
盖钧镒. 植物数量性状遗传体系. 科学出版社, 2003. pp 224-265, 351-370Gai JY. Genetic System of Quantitative Traits in Plants. Beijing: Science Press, 2003. pp 224-265, 351-370(in Chinese)[本文引用:2]
[16]
李余生, 朱镇, 张亚东, 赵凌, 王才林. 水稻稻曲病抗性的主基因+多基因混合遗传模型分析. 作物学报, 2008, 34: 1728-1733Li YS, ZhuZ, Zhang YD, ZhaoL, Wang CL. Genetic analysis of rice false smut resistance using major gene plus polygene mixed genetic model. Acta Agron Sin, 2008, 34: 1728-1733 (in Chinese with English abstract)[本文引用:1]
[17]
周清元, 崔翠, 阴涛, 陈东亮, 张正圣, 李加纳. 甘蓝型油菜角果长度的主基因+多基因混合遗传模型. 作物学报, 2014, 40: 1493-1500Zhou QY, CuiC, YinT, Chen DL, Zhang ZS, Li JN. Genetic analysis of silique length using mixture model of major gene plus polygene in Brassica napus L. Acta Agron Sin, 2014, 40: 1493-1500 (in Chinese with English abstract)[本文引用:1]
[18]
金岩, 吕艳艳, 付三雄, 戚存扣. 甘蓝型油菜苗期耐淹性状主基因+多基因遗传分析. 作物学报, 2014, 40: 1964-1972JinY, Lü YY, Fu SX, Qi CK. Inheritance of major gene plus polygene of water-logging tolerance in Brassica napus L. Acta Agron Sin, 2014, 40: 1964-1972 (in Chinese with English abstract)[本文引用:1]
[19]
田露申, 牛应泽, 余青青, 郭世星, 柳丽. 甘蓝型油菜白花性状的主基因+多基因遗传分析. 中国农业科学, 2009, 42: 3987-3995Tian LS, Niu YZ, Yu QQ, Guo SX, LiuL. Genetic analysis of white flower color with mixed model of major gene plus polygene in Brassica napus L. Sci Agric Sin, 2009, 42: 3987-3995 (in Chinese with English abstract)[本文引用:1]
[20]
顾慧, 戚存扣. 甘蓝型油菜(Brassica napus L. )抗倒伏性状的主基因+多基因遗传分析. 作物学报, 2008, 34: 376-381GuH, Qi CK. Genetic analysis of lodging resistance with mixed model of major gene plus polygene in Brassica napus L. Acta Agron Sin, 2008, 34: 376-381 (in Chinese with English abstract)[本文引用:1]
[21]
王会, 桑世飞, 梅德圣, 李云昌, 刘佳, 付丽, 王军, 陈玉峰, 胡琼. 甘蓝型油菜DH群体抗裂角性遗传分析. 中国油料作物学报, 2014, 36: 437-449WangH, Sang SF, Mei DS, Li YC, LiuJ, FuL, WangJ, Chen YF, HuQ. Inheritance analysis of pod shatter resistance using a DH population in Brassica napus L. Chin J Oil Crop Sci, 2014, 36: 437-449 (in Chinese with English abstract)[本文引用:1]
[22]
汪文祥, 胡琼, 梅德圣, 李云昌, 王会, 王军, 付丽, 刘佳. 基于图像处理的油菜分枝及角果着生角度测量方法. 中国油料作物学报, 2015, 37: 566-570Wang WX, HuQ, Mei DS, Li YC, WangH, WangJ, FuL, LiuJ. Evaluation of branch and pod angle measurement based on digital images from Brassica napus L. Chin J Oil Crop Sci, 2015, 37: 566-570 (in Chinese with English abstract)[本文引用:2]
[23]
曹锡文, 刘兵, 章元明. 植物数量性状分离分析Windows软件包SEA的研制. 南京农业大学学报, 2013, 36(6): 1-6Cao XW, LiuB, Zhang YM. SEA: a software package of segregation analysis of quantitative traits in plants. J Nanjing Agric Univ, 2013, 36(6): 1-6 (in Chinese with English abstract)[本文引用:1]
[24]
DiepenbrockW. Yield analysis of winter oilseed rape (Brassica napus L. ): a review. Field Crops Res, 2000, 67: 35-49[本文引用:1]
[25]
马霓, 张春雷, 李俊, 李光明. 种植密度对直播油菜结实期源库关系及产量的调节. 中国油料作物学报, 2009, 31: 180-184MaN, Zhang CL, LiJ, Li GM. Regulation of planting density on source-sink relationship and yield at seed-set stage of rapeseed (Brassica napus L. ). Chin J Oil Crop Sci, 2009, 31: 180-184 (in Chinese with English abstract)[本文引用:1]
[26]
朱珊, 李银水, 余常兵, 谢立华, 胡小加, 张树杰, 廖星, 廖祥生, 车志. 密度和氮肥用量对油菜产量及氮肥利用率的影响. 中国油料作物学报, 2013, 35: 179-184ZhuS, Li YS, Yu CB, Xie LH, Hu XJ, Zhang SJ, LiaoX, Liao XS, CheZ. Effects of planting density and nitrogen application rate on rapeseed yield and nitrogen efficiency. Chin J Oil Crop Sci, 2013, 35: 178-184 (in Chinese with English abstract)[本文引用:1]
[27]
马霓, 李云昌, 胡琼, 李俊, 李光明, 张春雷. 我国南方冬油菜机械化生产农艺研究进展. 中国油料作物学报, 2010, 32: 451-456MaN, Li YC, HuQ, LiJ, Li GM, Zhang CL. Research of agronomic techniques form mechanical production of winter rape (Brassica napus L. ). Chin J Oil Crop Sci, 2010, 32: 451-456 (in Chinese with English abstract)[本文引用:1]
[28]
王玫, 陈洪伟, 王红利, 刘克锋. 独脚金内酯调控植物分枝的研究进展. 园艺学报, 2014, 41: 1924-1934WangM, Chen HW, Wang HL, Liu KF. Research progress in regulatory role of strigolactones in shoot branching. Acta Hort Sin, 2014, 41: 1924-1934 (in Chinese with English abstract)[本文引用:1]
付三雄, 戚存扣. 甘蓝型油菜含油量的主基因+多基因遗传分析. 江苏农业学报, 2009, 25: 731-736Fu SX, Qi CK. Major gene plus polygene inheritance of oil content in Brassica napus L. Jiangsu J Agirc Sci, 2009, 25: 731-736 (in Chinese with English abstract)[本文引用:1]
 , 胡琼, 梅德圣, 李云昌, 周日金, 王会, 成洪涛, 付丽, 刘佳
, 胡琼, 梅德圣, 李云昌, 周日金, 王会, 成洪涛, 付丽, 刘佳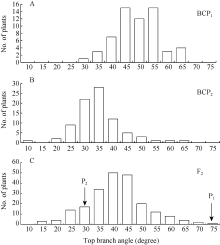



{kind=link}
{kind=link}