关键词:小麦; MYB转录因子; 防御反应; 小麦根腐病; 转基因 Development and Characterization of TaMYB86-Overexpressing Transgenic Wheat Lines with Resistance to Common Root Rot SHAN Tian-Lei, HONG Yan-Tao, DU Li-Pu, XU Hui-Jun, WEI Xue-Ning, ZHANG Zeng-Yan* National Key Facility for Crop Gene Resources and Genetic Improvement / Key Laboratory of Biology and Genetic Improvement of Triticeae Crops, Ministry of Agriculture / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:The study was supported by the National Major Project for Developing New GM Crops (2016ZX08002-001-004) and the National Natural Science Foundation of China (31471494) AbstractWheat common root rot, mainly caused by Bipolaris sorokiniana, is a difficultly prevent soil-borne disease of wheat ( Triticum aestivum L.) worldwide. TaMYB86, a B. sorokiniana-induced wheat MYB gene ,encodes a MYB transcription factor. We constructed the TaMYB86 overexpression vector pUbi:MYC-TaMYB86 and introduced TaMYB86 into Yangmai 16 via the particle bombardment. The TaMYB86 transgenic wheat lines on generations of T0-T3 were underwent by molecular characteristics analysis and disease resistance evaluation. The PCR and quantitative RT-PCR results showed that the alien TaMYB86was introduced into three transgenic wheat lines, and the relative transcriptional level of TaMYB86was apparently higher in transgenic wheat lines than in non-transformed Yangmai 16. As Western blot results presented, the introduced MYC-TaMYB86 gene was translated into the MYC-TaMYB86 protein in the three overexpressing transgenic lines, but not in non-transformed Yangmai 16. The infection types and disease indexes of three TaMYB86 transgenic wheat lines were significantly lower than those of non-transformation Yangmai 16 ( t-test, P< 0.01). The transcript levels of 3 wheat defense genes ( PR10, PR17c, and Chit1) were significantly elevated in three transgenic wheat lines than in the non-transformed Yangmai 16. These results indicate that overexpression of TaMYB86 enhances significantly resistance to B. sorokiniana in transgenic wheat lines and TaMYB86 plays a positive role in defense response to B. sorokiniana.
Keyword: Triticum aestivum; MYB transcription factor; Defense response; Common root rot; Transgene Show Figures Show Figures
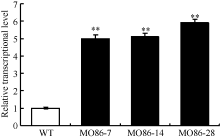
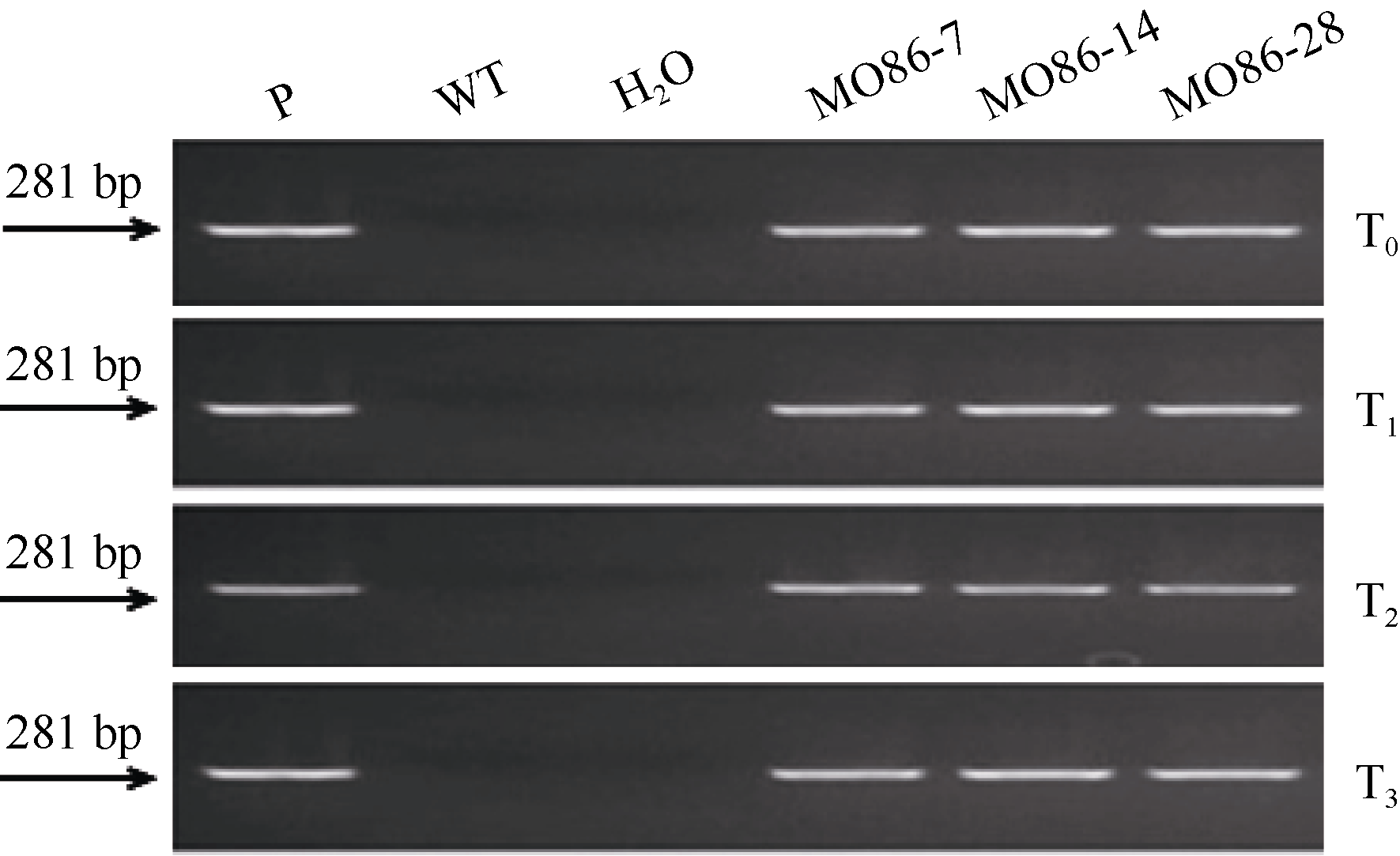
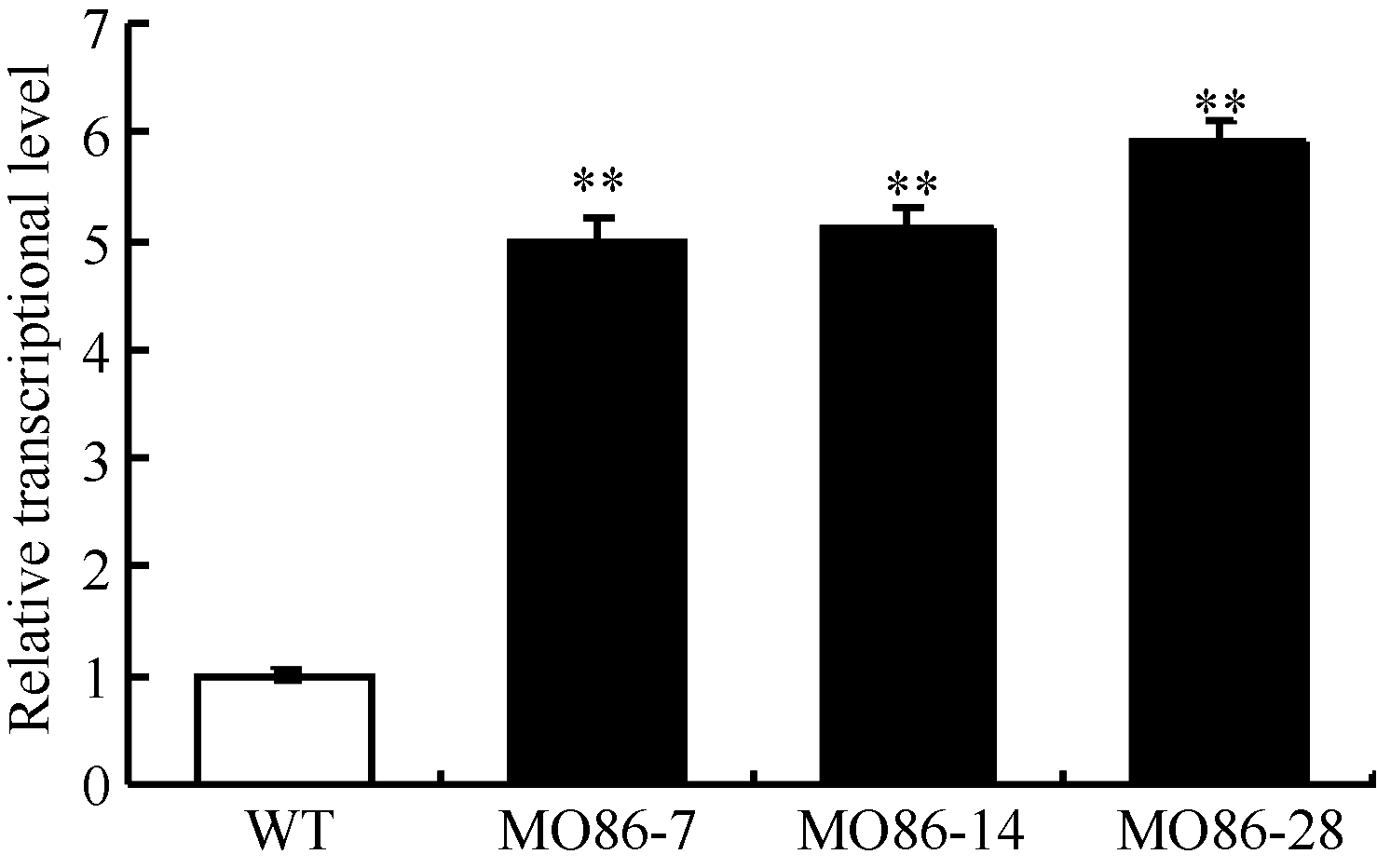
图3 过表达株系中TaMYB86的转录水平分析WT: 未转基因扬麦16; MO86-7、MO86-14、MO86-28: TaMYB86过表达株系。每个样本进行3次重复实验, 采用t检验分析差异显著性; * * P < 0.01概率水平极显著。Fig. 3 qRT-PCR analysis of TaMYB86transcriptional levels in transgenic wheat linesWT: wild type, non-transformed Yangmai 16; MO86-7, MO86-14, MO86-28: TaMYB86transgenic wheat lines. Three biological replicates for each line were averaged and statistically treated (t-test; * * P < 0.01).
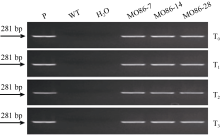
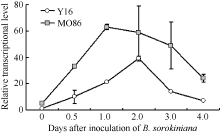
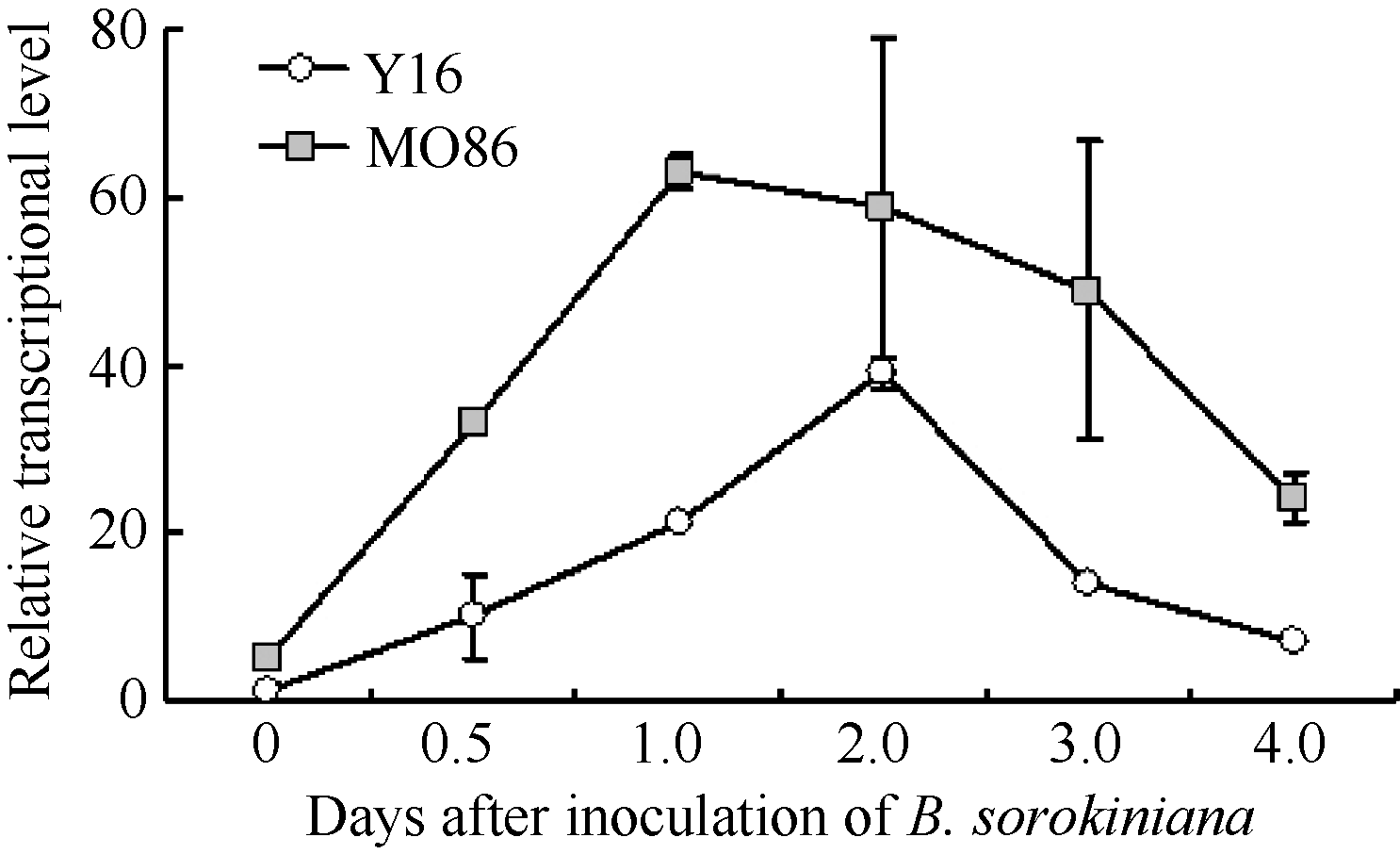
图4 根腐病菌接种后TaMYB86的表达模式Y16: 未转基因扬麦16; MO86: TaMYB86过表达株系MO86-7、MO86-14、MO86-28的混合样品。每个样本进行3次重复实验。Fig. 4 Transcriptional patterns of TaMYB86 in B. sorokiniana-inoculated wheatY16: non-transformed Yangmai 16; MO86: pooled samples of transgenic wheat lines MO86-7, MO86-14, and MO86-28.
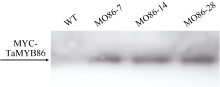
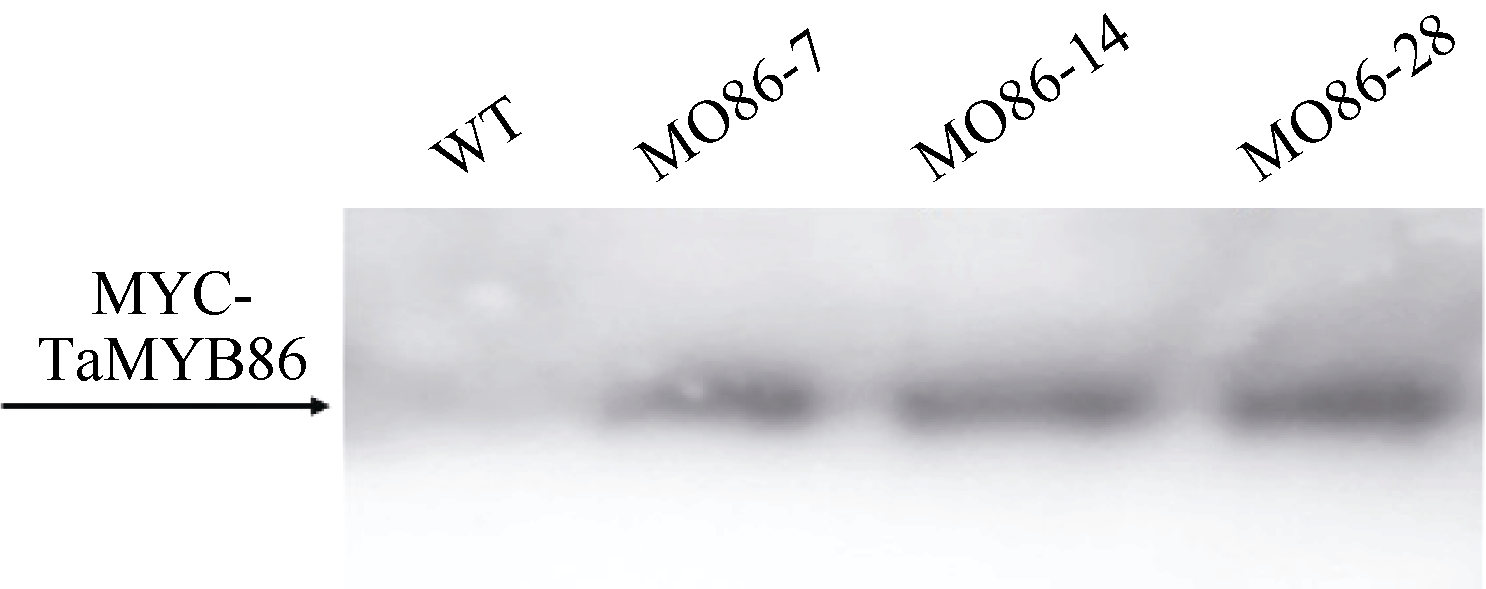
图5 转TaMYB86基因过表达株系和未转基因扬麦16的Western杂交分析WT: 未转基因扬麦16; MO86-7, MO86-14, MO86-28: TaMYB86过表达株系。箭头指示MYC-TaMYB86融合蛋白的位置。Fig. 5 Western blot pattern of TaMYB86-overexpressed transgenic lines and non-transformed Yangmai 16 using an anti-MYC antibodyWT: wild type, non-transformed Yangmai 16; MO86-7, MO86-14, MO86-28: TaMYB86transgenic wheat lines. The arrow shows the position of MYC-TaMYB86 fusion protein.
表1 Table 1 表1(Table 1)
表1 转TaMYB86基因过表达株系与未转基因扬麦16的根腐病抗性鉴定 Table 1 Common root rot response of TaMYB86-overexpressed transgenic lines and non-transformed wheat Yangmai 16
株系 Line
T1
T2
T3
IT
DI
IT
DI
IT
DI
MO86-7
1.27± 0.43* *
31.67± 10.87* *
1.51± 0.45* *
37.76± 11.33* *
1.67± 0.49* *
41.67± 12.13* *
MO86-14
2.00± 0* *
50.00± 0* *
1.50± 0.53* *
37.50± 13.18* *
1.25± 0.43* *
31.25± 10.69* *
MO86-28
1.80± 0.45* *
45.00± 11.18* *
1.54± 0.34* *
38.43± 8.42* *
1.50± 0.50* *
37.50± 12.50* *
CI12633
1.15± 0.23
28.69± 5.77
1.07± 0.09
26.85± 2.21
1.17± 0.24
29.33± 6.10
阴性植株 NTP
2.80± 0.24
70.00± 6.12
2.15± 0.28
53.75± 7.07
2.56± 0.66
64.00± 16.55
扬麦16 Yangmai 16
3.00± 0.17
75.04± 4.14
2.17± 0.46
54.17± 11.49
2.62± 0.77
65.38± 19.20
MO 86-7, MO86-14, and MO86-28 represent TaMYB86-overexpression transgenic wheat lines. Negative-transgenic plants indicate negative PCR plants. IT represents infection type. DI indicates disease index of common root rot. Response value for each line is the mean of at least 10 plants in each generation. Statistically significant differences between each line ofTaMYB86-overexpression wheat and non-transformed Yangmai 16 were determined using t-test (* * P < 0.01). NTP: negative-transgenic plants. MO86-7、MO86-14、MO86-28代表TaMYB86-过表达的转基因株系; 阴性植株表示转基因检测为阴性的植株; 扬麦16代表非转基因扬麦16。IT: 感染类型; DI: 病情指数; 每个株系对至少10株小麦进行抗病鉴定; 采用t检验分析差异显著性; * * P < 0.01概率水平极显著。
表1 转TaMYB86基因过表达株系与未转基因扬麦16的根腐病抗性鉴定 Table 1 Common root rot response of TaMYB86-overexpressed transgenic lines and non-transformed wheat Yangmai 16
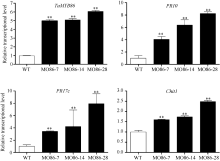
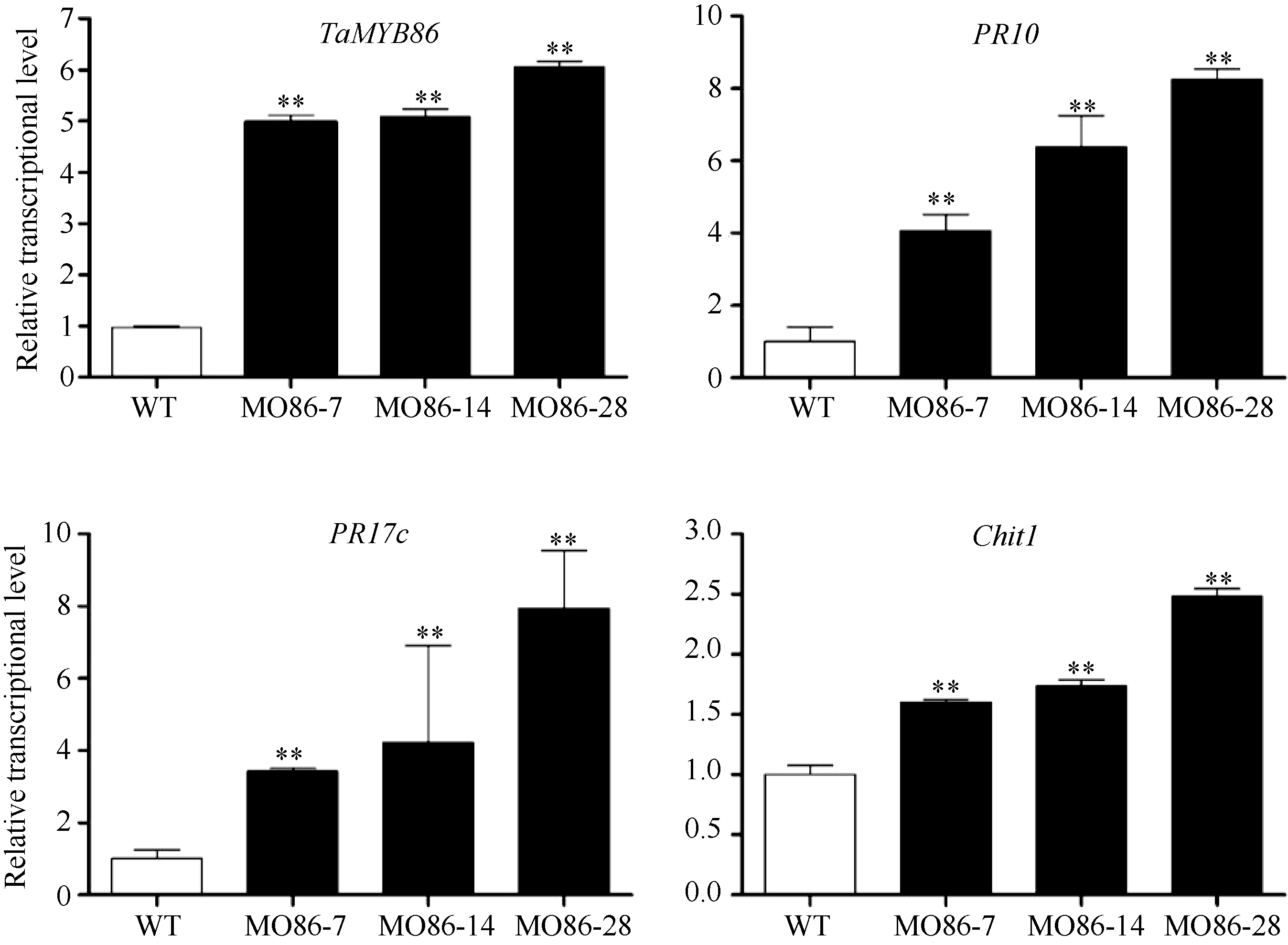
图6TaMYB86过表达株系中TaMYB86和下游防卫基因的qRT-PCR分析接种根腐病菌47 d后TaMYB86过表达株系和未转基因扬麦16 (WT)植株中TaMYB86和下游防卫基因的转录表达。防卫基因包括 PR10 (CA613496)、PR17c (TA65181)和Chit1 (CA665185)。采用t检验分析差异显著性; * * P < 0.01极显著。Fig. 6 qRT-PCR analysis of the relative transcript level of TaMYB86 and three defense genes in TaMYB86-overexpressing transgenic linesThe expression of TaMYB86 and three defense genes in TaMYB86-overexpressing transgenic and non-transformed wheat Yangmai 16 (WT) inoculated with B. sorokiniana for 47 d. Defense genes include PR10 (CA613496), PR17c (TA65181) and Chit1 (CA665185). Three biological replicates of each line were averaged and statistically treated (t-test; * * P < 0.01).
4 结论通过基因枪转化、分子检测和抗根腐病鉴定, 创制、筛选出抗根腐病的TaMYB86过表达转基因小麦新种质3份, 证明TaMYB86在小麦防御根腐病过程中起正向调控作用。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
KumarJ, SchäferP, HeckelhovenR, LangenG, BaltruschatH, SteinE, NagarajanS, Kogel KH. Bipolaris sorokiniana, a cereal pathogen of global concern: cytological and molecular approaches towards better control. Mol Plant Pathol, 2002, 3(4): 185-195[本文引用:1]
[2]
贾廷祥, 吴桂本, 刘传德. 我国小麦根腐性病害研究现状及防治对策. 中国农业科学, 1995, 28: 41-48Jia YX, Wu GB, Liu CD. The present research situation and control countermeasure of root rots in wheat. Sci Agric Sin, 1995, 28: 41-48 (in Chinese with English abstract)[本文引用:1]
[3]
Li HJ, Conner RL, ChenQ, Li HY, LarocheA, Graf RJ, Kuzyk AD. The transfer and characterization of resistance to common root rot from Thinopyrum ponticum to wheat. Genome, 2004, 47: 215-223[本文引用:2]
HagaN, KatoK, MuraseM, ArakiS, KuboM, DemuraT, SuzukiK, MüllerI, VossU, Jürgens G. ItoM. R1R2R3-Myb proteins positively regulate cytokinesis through activation of KNOLLE transcription in Arabidopsis thaliana. Development, 2007, 134: 1101-1110[本文引用:1]
[6]
ZhouJ, LeeC, ZhongR, Ye ZH. MYB58 and MYB63 are transcriptional activators of the lignin biosynthetic pathway during secondary cell wall formation in Arabidopsis. Plant Cell, 2009, 21: 248-266[本文引用:1]
[7]
Millar AA, GublerF. The Arabidopsis GAMYB-like genes, MYB33 and MYB65, are microRNA-regulated genes that redundantly facilitate anther development. Plant Cell, 2005, 17: 705-721[本文引用:1]
[8]
MullerD, SchmitzG, TheresK. Blind homologous R2R3 Myb genes control the pattern of lateral meristem initiation in Arabidopsis. Plant Cell, 2006, 18: 586-597[本文引用:1]
[9]
AbeH, UraoT, ItoT, SekiM, ShinozakiK, Yamaguchi- ShinozakiK. Arabidopsis AtMYC2 (bHLH) and AtMYB2 (MYB) function as transcriptional activators in abscisic acid signaling. Plant Cell, 2003, 15: 63-78[本文引用:1]
[10]
JungC, Seo JS, Han SW, Koo YJ, Kim CH, Song SI, Nahm BH, Choi YD, Cheong JJ. Overexpression of AtMYB44 enhances stomatal closure to confer abiotic stress tolerance in transgenic Arabidopsis. Plant Physiol, 2008, 146: 623-635[本文引用:1]
[11]
Dai XY, Xu YY, Ma QB, Xu WY, WangT, Xue YB, ChongK. Overexpression of an R1R2R3 MYB gene, OsMYB3R-2, increases tolerance to freezing, drought, and salt stress in transgenic Arabidopsis. Plant Physiol, 2007, 143: 1739-1751[本文引用:1]
[12]
Su CF, Wang YC, Hsieh TH, Lu CA, Tseng TH, Yu S M. A novel MYBS3-dependent pathway confers cold tolerance in rice. Plant Physiol, 2010, 153: 145-158[本文引用:1]
[13]
MengisteT, ChenX, SalmeronJ, DietrichR. The BOTRYTIS SUSCEPTIBLE1 gene encodes an R2R3MYB transcription factor protein that is required for biotic and abiotic stress responses in Arabidopsis. Plant Cell, 2003, 15: 2551-2565[本文引用:2]
[14]
Seo PJ, XiangF, QiaoM, Park JY, Lee YN, Kim SG, Lee YH, Park WJ, Park CM. The MYB96 transcription factor mediates abscisic acid signaling during drought stress response in Arabidopsis. Plant Physiol, 2009, 151: 275-289[本文引用:1]
[15]
Seo PJ, Park CM. MYB96-mediated abscisic acid signals induce pathogen resistance response by promoting salicylic acid biosynthesis in Arabidopsis. New Phytol, 2010, 186: 471-483[本文引用:1]
LiuX, Yang LH, Zhou XY, Zhou MP, LuY, Ma LJ, Zhang ZY. Transgenic wheat expressing Thinopyrum intermedium MYB transcription factor TiMYB2R-1 shows enhanced resistance to the take-all disease. J Exp Bot, 2013, 64: 2243-2253[本文引用:1]
[18]
Al-Attala MN, WangX, Abou-Attia MA, DuanX, KangZ. A novel TaMYB4 transcription factor involved in the defence response against Puccinia striiformis f. sp. tritici and abiotic stresses. Plant Mol Biol, 2014, 84: 589-603[本文引用:1]
[19]
Zhang ZJ, Chen JM, Su YY, Liu HM, Chen YE, Luo PG, Du XG, WangD, Zhang HY. TaLHY, a 1R-MYB transcription factor, plays an important role in disease resistance against stripe rust fungus and ear heading in wheat. PLoS One, 2015, 10, doi: DOI:10.1371/journal.pone.0127723[本文引用:1]
[20]
Christensen AH, Quail PH. Ubiquitin promoter-based vectors for high-level expression of selectable and /or screenable marker genes in monocotyledonous plants. Transgenic Res, 1996, 5: 213-218[本文引用:1]
[21]
Zhu XL, YangK, Wei XN, Zhang QF, RongW, Du LP, Ye XG, QiL, Zhang ZY. The wheat AGC kinase TaAGC1 is a positive contributor to host resistance to the necrotrophic pathogen Rhizoctonia cerealis. J Exp Bot, 2015, 66: 6591-6603[本文引用:1]
[22]
徐惠君, 庞俊兰, 叶兴国, 杜丽璞, 李连城, 辛志勇, 马有志, 陈剑平, 陈炯, 程顺和, 吴宏亚. 基因枪介导法向小麦导入黄花叶病毒复制酶基因的研究. 作物学报, 2001, 27: 688-693Xu HJ, Pang JL, Ye XG, Du LP, Li LC, Xin ZY, Ma YZ, Chen JP, ChenJ, Cheng SH, Wu HY. Study on the gene transferring of Nib8 into wheat for its resistance to the Yellow mosaic virus by bombardment. Acta Agron Sin, 2001, 27: 688-694 (in Chinese with English abstract)[本文引用:1]
[23]
Sharp PJ, KreisM, Shewry PR, Gale MD. Location of β-amylase sequences in wheat and its relatives. Theor Appl Genet, 1988, 75: 286-290[本文引用:1]
[24]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25: 402-408[本文引用:1]
[25]
DongN, LiuX, LuY, Du LP, Xu HJ, Liu HX, Xin ZY, Zhang ZY. Overexpression of TaPIEP1, a pathogen-induced ERF gene of wheat, confers host-enhanced resistance to fungal pathogen Bipolaris sorokiniana. Funct Integr Genomic, 2010, 10: 215-226[本文引用:2]
[26]
DanielX, LacommeC, Morel JB, RobyD. A novel myb oncogene homologue in Arabidopsis thaliana related to hypersensitive cell death. Plant J, 1999, 20: 57-66[本文引用:1]
[27]
VailleauF, DanielX, TronchetM, Montillet JL, TriantaphylidesC, RobyD. A R2R3-MYB gene, AtMYB30, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack. Proc Natl Acad Sci USA, 2002, 99: 10179-10184[本文引用:1]
[28]
ZhangH, ZhangD, ChenJ, YangY, HuangZ, HuangD, Wang XC, HuangR. Tomato stress-responsive factor TSRF1 interacts with ethylene responsive element GCC box and regulates pathogen resistance to Ralstonia solanacearum. Plant Mol Biol, 2004, 55: 825-834[本文引用:1]
[29]
ChenL, Zhang ZY, Liang HX, Liu HX, Du LP, Xu HJ, Xin ZY. Overexpression of TiERF1 enhances resistance to sharp eyespot in transgenic wheat. J Exp Bot, 2008, 59: 4195-4204[本文引用:1]
[30]
Zhang ZY, LiuX, Wang XD, Zhou MP, Zhou XY, Ye XG, Wei XN. An R2R3 MYB transcription factor in wheat, TaPIMP1, mediates host resistance to Bipolaris sorokiniana and drought stresses through regulation of defense- and stress-related genes. New Phytol, 2012, 196: 1155-1170[本文引用:1]
[31]
LiuX, Yang LH, Zhou XY, Zhou MP, LuY, Ma LJ, Ma HX, Zhang ZY. Transgenic wheat expressing Thinopyrum intermedium MYB transcription factor TiMYB2R-1 shows enhanced resistance to the take-all disease. J Exp Bot, 2013, 64: 2243-2253[本文引用:1]
[32]
Zhu XL, QiL, LiuX, Cai SB, Xu HJ, Huang RF, Li JR, Wei XN, Zhang ZY. The wheat ethylene response factor transcription factor pathogen-induced ERF1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiol, 2014, 164: 1499-1514[本文引用:1]
[33]
ShinS, Mackintosh CA, LewisJ, Heinen SJ, RadmerL, Dill-MackyR, Baldridge GD, Zeyen RJ, Muehlbauer GJ. Transgenic wheat expressing a barley class II chitinase gene has enhanced resistance against Fusarium graminearum. J Exp Bot, 2008, 59: 2371-2378[本文引用:1]
[34]
LiZ, Zhou MP, Zhang ZY, Ren LJ, Du LP, Zhang BQ, Xu HJ, Xin ZY. Expression of a radish defensin in transgenic wheat confers increased resistance to Fusarium graminearum and Rhizoctonia cerealis. Funct Integr Genomics, 2011, 11: 63-70[本文引用:1]
[35]
Zhu XL, LiZ, Xu HJ, Zhou MP, Du LP, Zhang ZY. Overexpression of wheat lipid transfer protein gene TaLTP5 increases resistances to Cochliobolus sativus and Fusarium graminearum in transgenic wheat. Funct Integr Genomics, 2012, 12: 481-488[本文引用:1]


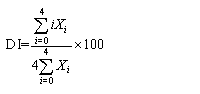

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}