关键词:油菜; 收获指数; 相关分析; 通径分析 Analysis of Yield Components with High Harvest Index in Brassica napus under Environments Fitting Different Yield Levels LU Kun1,**, SHEN Ge-Zi1,**, LIANG Ying1, FU Ming-Lian2, HE Bin2, TIE Lin-Mei3, ZHANG Ye1, PENG Liu1, LI Jia-Na1,* 1College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China
2 Industrial Crops Institute, Yunnan Academy of Agricultural Sciences, Kunmimg 650205, China
3 Agricultural Technology Extension Station in Lincang City, Lincang 677000, China
Fund:This study was supported by the National Science Foundation of China (U1302266 and 31571701), the National Basic Research Program of China (2015CB150201), the 111 Project of Chinese Ministry of Education (B12006), and the China Agriculture Research System (CARS-13) AbstractLow harvest index is a bottleneck for further improvement of seed yield and oil production in rapeseed ( Brassica napus). A total of 321 B. napus accessions with normal growth and maturation at both the super high yield production area Lincang, Yunnan province and the major production area of the upper Yangtze River Basin, Beibei, Chongqing were chosen in this study. Variance of yield-related traits and their differences between two locations were compared, and the relationship between yield harvest index (YHI), oil production harvest index (OHI) and 17 yield component traits were also investigated by correlation and path analyses. At Yunnan, the main reasons for high yield of B. napus were adequate illumination, big diurnal temperature difference, sufficient photosynthate, more biomass, higher number of siliques per plant and seed number per silique, especially the number of siliques on the secondary branches, which might play key roles in improvement of B. napus yield. The silique number on the main inflorescence, primary and secondary branches was significantly and positively correlated with YHI at Chongqing, but the reverse relationship was observed between YHI and silique number on main inflorescence at Yunnan. The number of seeds per silique on the main inflorescence, primary and secondary branches at Yunnan were significantly higher than those at Chongqing, and the number of seeds per silique on the main inflorescence and primary branch showed significant or extremely significantly positive correlation with YHI and OHI simultaneously at both two cultivated areas, suggesting that sufficient accumulation of photosynthate and high grain filling efficiency are necessary for large number of seeds per silique. Thousand-seed weights on the main inflorescence, primary and secondary branches at Yunnan were all lower than those at Chongqing, suggesting that once the photosynthate is insufficient, the seed number per plant will decrease and partial seeds will become the priority for seed-filling in B. napus, to ensure the quality of offspring seeds. Under the high yield production condition of Yunnan, thousand-seed weights on three different parts showed significant or remarkably significantly positive correlation with YHI and OHI, while those at Chongqing were not, indicating that thousand-seed weight is determined by the transport capacity of photosynthate under the condition of sufficient illumination. In summary, the number of siliques on the main inflorescence and seed yield per plant are two key factors for increasing YHI under adequate illumination environment. But the composition of YHI is more complicated under weak illumination environment. Thus, higher YHI could be achieved when the yield components on the main inflorescence and primary branch are organic ally integrated and biological yield is restrained.
Keyword: Brassica napus; Harvest index; Correlation analysis; Path analysis Show Figures Show Figures
作物产量提高主要有两种途径, 一是在生物学产量不变的情况下提高收获指数(harvest index); 二是在收获指数一定的情况下提高生物学产量[1, 2]。众多研究表明, 近年来世界水稻(Oryza sativa)、小麦(Triticum aestivum)和大麦(Hordeum vulgare)等主要作物产量的增长主要得益于收获指数的改良, 收获指数已成为评价作物品种选育效果的重要内容[3, 4, 5]。相当多的****研究了水稻、小麦收获指数的遗传、改良方式和在育种中的应用[6, 7, 8, 9, 10], 但大部分****仍集中在对作物产量的研究上。 甘蓝型油菜(Brassica napus)是高油分的油料作物和高蛋白的饲料作物, 也是潜在的能源作物。培育高产优质新品种一直是油菜育种的首要目标。与其他作物相比, 油菜的生物产量很高, 但收获指数显著偏低[5], 因此提高收获指数是实现油菜高产的有效途径。到目前为止, 关于油菜收获指数的系统研究较少。黄露等[11]对四川29个油菜杂交种的收获指数、生物产量、经济产量、粒壳重比、粒枝重比、粒茎重比等性状的相关分析发现, 粒茎重比对收获指数影响最大, 粒枝重比次之, 粒壳重比最小, 在生物产量相当的情况下, 适当控制茎秆重有利于油菜产量的提高。袁婺洲等[12]研究发现甘蓝型油菜角果皮内的淀粉酶活性, 可溶性糖含量及32P向籽粒的运输强度等均与油菜的收获指数呈显著正相关, 而角果皮内的淀粉含量则与收获指数呈负相关, 油菜粒壳比, 角杆比及千粒重与收获指数呈正相关, 杆重与收获指数呈负相关[13]。Luo等[14]利用155份材料的SNPs芯片数据, 对油菜株高、分枝数、生物产量、籽粒产量和收获指数进行了全基因组关联分析, 在C亚基因组上找到9个与收获指数紧密关联的SNPs标记, 并发现株高与收获指数呈显著负相关。 云南临沧独特的生态环境有利于油菜高产, 主推品种云油杂2号大面积单产可达4500 kg hm-2以上, 远高于长江流域油菜主产区。经前期调查, 云南临沧高产油菜的产量构成特点主要为有效分枝、单株角果和每角粒数多, 特别是二次有效分枝和二次分枝角果多、籽粒多且饱满。因此, 研究相同油菜品种(系)在高产环境(云南临沧)和一般环境(重庆北碚)下各产量组成性状差异和不同环境下收获指数与各产量组成性状的相关性, 可解析收获指数及构成因素受基因型和环境影响的规律与原因, 为油菜高收获指数育种提供理论依据。 1 材料与方法1.1 材料与试验设计从国内外广泛收集各种资源材料600多份, 通过预备试验, 选择在云南和重庆均能正常生长和成熟的321份甘蓝型油菜品种, 于2013年9月至2014年5月分别在重庆市油菜工程技术研究中心歇马实验种植基地(29º 45'39.99" N, 106º 22'38.47" E, 海拔238.57 m)和云南省临沧市农业技术推广站临翔区博尚镇勐准村实验种植基地(23º 43'56.69" N, 100º 02'4.79" E, 海拔1819.50 m), 采用育苗移栽方式, 参照卢坤等[15]的方法按完全随机区组设计三重复同时种植, 每小区2行, 每行15株, 行距40 cm, 株距20 cm。田间管理同常规生产, 确保同一地区所有样本的生长环境一致。 1.2 测定项目及方法1.2.1 产量组成性状 成熟期从每小区选择生长正常、无病害、无机械损伤的10个样株, 分别调查每株生物学产量(biomass per plant, PB)、主序角果数(number of effective siliques on the main inflorescence, MIS)、一次有效分枝角果数(number of effective siliques on the primary branch, PBS, 简称一次角果数)和二次有效分枝角果数(number of effective siliques on the secondary branch, SBS, 简称二次角果数)。然后按主序、一次有效分枝、二次有效分枝分别装袋, 自然风干后考察主序粒重(seed weight on the main inflorescence, SWMI)、一次有效分枝粒重(seed weight on the primary branch, SWPB, 简称一次粒重)、二次有效分枝粒重(seed weight on the secondary branch, SWSB, 简称二次粒重)和3个部位的千粒重, 分别标识为TSWMI (thousand-seed weight on the main inflorescence)、TSWPB (thousand- seed weight on the primary branch)、TSWSB (thousand-seed weight on the secondary branch); 单株产量(seed yield per plant, PY)为主序粒重、一次粒重和二次粒重之和。最后, 分别计算主序每角粒数(number of seeds per silique on the main inflorescence, SSMI)、一次有效分枝每角粒数(number of seeds per silique on the primary branch, SSPB, 简称一次每角粒数)、二次有效分枝每角粒数(number of seeds per silique on the secondary branch, SSSB, 简称二次每角粒数), 不同部位的每角粒数计算公式为(粒重/千粒重× 1000)/角果数。根据上述性状考察结果, 计算单株角果数(silique number per plant, SP)、单株平均每角粒数(mean seed number per silique, MSP)和单株平均千粒重(mean thousand-seed weight per plant, MTSWP)。 1.2.2 产量和产油量收获指数 成熟期从每小区另外收获5株正常植株, 将所有分枝齐茎秆处剪断装袋, 晒干后称重作为上部干重; 再将所有角果脱粒, 种子晒干后获得单株经济产量; 使用NIRSystems.5000近红外分析仪测定各材料的含油量(oil content, OC); 将5株茎杆齐子叶节切断, 晒干后称重得到单株平均茎杆干重; 将上部干重与茎杆干重相加得到全株生物产量。按下述公式计算各材料的产量收获指数和产油量收获指数。产量收获指数(yield harvest index, YHI) = 经济产量/生物产量 = 5株种子总重/(5株上部干重+5株茎秆干重); 产油量收获指数(oil production harvest index, OHI) = 产油量/生物产量=含油量× 产量/生物产量=含油量× 产量收获指数。 1.3 气象数据获取材料种植期间两地的温度、降雨、湿度、风速和光照时长数据来自美国国家海洋与大气管理局(NOAA)地球系统研究实验室物理科学部(http:// www.esrl.noaa.gov/psd/), 并以ncBrowse工具从NetCDF文件中获取相关气象数据。太阳辐射数据来自美国国家航空航天局(NASA)大气科学数据中心(https://eosweb.larc.nasa.gov/), 为两地22年(1983— 2005年)的月平均数。 1.4 数据分析利用Microsoft Excel 2010进行性状数据描述性统计分析和频率分布图制作。采用DPS7.05统计分析软件对两地各性状进行方差分析和Duncan’ s新复极差检验, 将方差组分分解为区组、环境、基因型、环境和基因型互作。用DPS7.05统计分析软件进行两地17个油菜产量组成性状与产量收获指数和产油量收获指数的相关分析和通径分析。
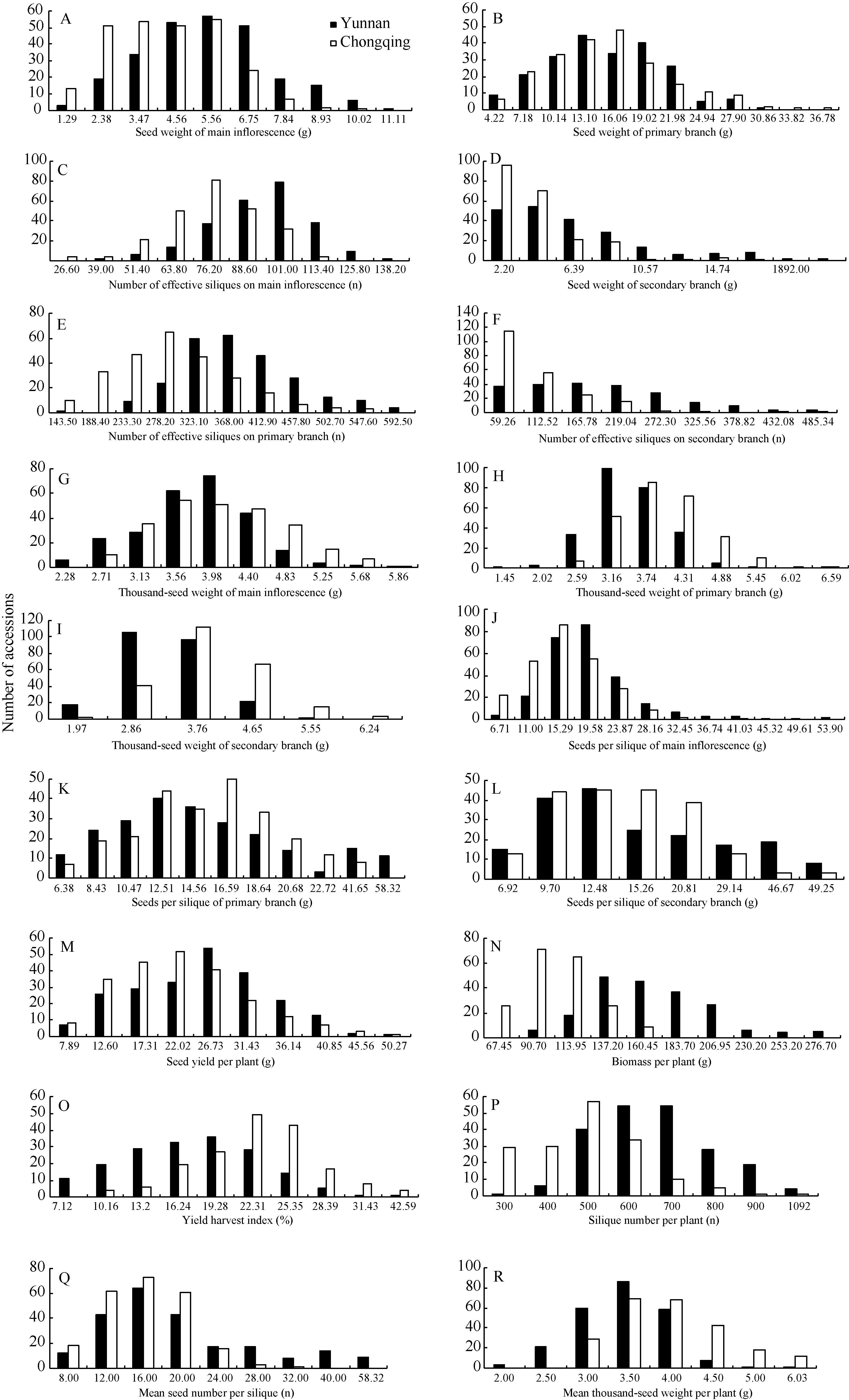
表1 重庆和云南321份材料20个性状的统计分析 Table 1 Statistical analysis of 20 traits in 321 B. napus accessions cultivated at Yunnan and Chongqing
性状 Trait
云南 Yunnan
重庆 Chongqing
均值的差值 D-value
均值± 标准差 Mean± SD
变幅 Range
变异系数 CV
均值± 标准差 Mean± SD
变幅 Range
变异系数 CV
产量收获指数 YHI
15.89± 5.60
4.08-42.59
35.22
21.18± 4.91
7.34-34.47
23.20
-5.29* *
产油量收获指数 OHI
8.05± 2.93
1.72-14.69
36.43
7.91± 3.07
3.48-22.40
38.76
0.14
单株经济产量 PY
23.13± 8.83
3.18-47.67
38.18
20.29± 8.33
3.95-50.27
41.09
2.84* *
单株生物产量 PB
153.67± 39.83
70.06-323.24
25.92
92.44± 23.35
44.23-157.21
25.26
61.23* *
单株角果数 SP
614.33± 138.36
296.60-1046.20
22.52
424.87± 129.90
182.00-1092.00
30.57
189.46* *
单株平均每角粒数 MSP
16.93± 7.28
5.00-58.32
43.04
13.58± 5.26
5.00-37.30
38.74
3.35* *
单株平均千粒重 MTSWP
3.20± 0.53
1.90-5.54
16.42
3.72± 0.66
2.57-6.03
17.76
-0.52
含油量 OC
41.21± 2.66
31.81-52.40
6.45
38.28± 2.09
33.44-44.73
5.46
2.93* *
主序粒重 SWMI
4.98± 1.89
0.55-11.11
37.91
3.74± 1.72
0.20-9.52
45.90
1.24* *
一次粒重 SWPB
13.59± 5.52
1.26-30.03
40.64
13.96± 6.02
1.94-36.78
43.14
-0.40
二次粒重 SWSB
5.38± 4.31
0.18-21.01
80.03
3.14± 2.60
0.12-14.81
82.95
2.24* *
主序角果数 MIS
87.55± 17.00
33.60-138.20
19.42
70.48± 16.58
14.20-107.40
23.52
17.07* *
一次角果数 PBS
354.20± 80.84
126.80-637.40
22.82
268.70± 82.03
98.60-538.00
30.53
85.50* *
二次角果数 SBS
165.64± 103.75
8.00-538.60
62.63
75.10± 64.76
6.00-468.80
86.22
90.54* *
主序千粒重 TSWMI
3.58± 0.65
1.86-5.86
18.11
3.88± 078
3.02-5.44
20.23
-0.30* *
一次千粒重 TSWPB
3.15± 0.61
0.88-6.31
19.35
3.71± 0.66
2.32-6.59
17.82
-0.56* *
二次千粒重 TSWSB
2.87± 0.64
1.07-5.15
22.32
3.55± 0.75
1.80-6.24
20.98
-0.68
主序每角粒数 SSMI
17.53± 6.86
5.18-53.90
39.13
13.98± 5.54
5.00-37.30
39.60
3.55* *
一次每角粒数 SSPB
17.95± 16.48
5.00-58.32
91.82
14.33± 4.51
5.00-31.00
31.47
3.62* *
二次每角粒数 SSSB
16.80± 15.97
5.00-49.25
95.04
12.84± 7.80
5.00-33.75
60.75
3.96* *
SD: standard deviation; CV: coefficient of variation; D-value: Difference of the mean value of a trait between Yunnan and Chongqing; YHI: yield harvest index (%); OHI: oil production harvest index of (%); PY: seed yield per plant (g); PB: biomass per plant (g); SP: silique number per plant (n); MSP: mean seed number per silique (n); MTSWP: mean thousand-seed weight per plant (g); OC: oil content (%); SWMI: seed weight on the main inflorescence (g); SWPB: seed weight on the primary branch (g); SWSB: seed weight on the secondary branch (g); MIS: number of effective siliques on the main inflorescence (n); PBS: number of effective siliques on the primary branch (n); SBS: number of effective siliques on the secondary branch (n); TSWMI: thousand-seed weight on the main inflorescence (g); TSWPB: thousand-seed weight on the primary branch (g); TSWSB: thousand-seed weight on the secondary branch (g); SSMI: number of seeds per silique on the main inflorescence (n); SSPB: number of seeds per silique on the primary branch (n); SSSB: number of seeds per silique on the secondary branch (n). * * indicates significance at the 0.01 level. SD: 标准差; CV: 变异系数; D-value: 云南和重庆两地均值的差值; YHI: 产量收获指数(%); OHI: 产油量收获指数(%); PY: 单株经济产量(g); PB: 单株生物产量(g); MSP: 单株角果数(n); MSSP: 单株平均每角粒数(n); MTSWP: 单株平均千粒重(g); OC: 含油量(%); SWMI: 主序粒重(g); SWPB: 一次粒重(g); SWSB: 二次粒重(g); MIS: 主序角果数(n); PBS: 一次角果数(n); SBS: 二次角果数(n); TSWMI: 主序千粒重(g); TSWPB: 一次千粒重(g); TSWSB: 二次千粒重(g); SSMI: 主序每角粒数(n); SSPB: 一次每角粒数(n); SSSB: 二次每角粒数(n); * * 表示在0.01水平差异显著。
表1 重庆和云南321份材料20个性状的统计分析 Table 1 Statistical analysis of 20 traits in 321 B. napus accessions cultivated at Yunnan and Chongqing
表2 Table 2 表2(Table 2)
表2 云南和重庆321份甘蓝型油菜20个性状的方差分析 Table 2 Analysis of variance of 20 traits in 321B. napusaccessions cultivated at Chongqing and Yunnan
变异来源 Source of variation
区组 Among blocks
环境 Environment (E)
基因 Genotype (G)
环境× 基因 E× G
产量收获指数YHI
1.78
90.38* *
1.02
3.85
产油量收获指数OHI
12.39* *
0.59
1.13
3.37* *
含油量OC
0.81
366.67* *
3.46* *
2.22* *
单株经济产量PY
5.35* *
11.74* *
0.90
3.93* *
单株生物产量PB
0.40
116.21* *
0.94
1.36* *
单株平均千粒重MTSWP
1.22
225.59* *
6.96* *
4.83* *
主序粒重SWMI
2.77*
74.71* *
1.46* *
2.21* *
一次粒重SWPB
3.02*
10.87
0.88
3.36* *
二次粒重 SWSB
1.72
42.27* *
1.01
3.09* *
单株角果数 SP
1.31
421.23* *
3.80* *
2.26* *
主序角果数MIS
1.52
160.97* *
1.49* *
3.74* *
一次角果数PBS
1.28
181.31* *
1.55* *
2.34* *
二次角果数SBS
0.53
140.51* *
1.37*
3.11* *
主序千粒重TSWMI
1.84
24.73* *
1.27*
4.36* *
一次千粒重TSWPB
1.49
119.70* *
1.44* *
3.72* *
二次千粒重TSWSB
1.82
109.76* *
1.15
2.28* *
单株平均每角粒数MSP
1.35
105.53* *
3.32* *
3.52* *
主序每角粒数SSMI
0.39
47.18* *
1.51* *
1.18*
一次每角粒数SSPB
0.86
17.11* *
0.97
4.32* *
二次每角粒数SSSB
1.35
10.87* *
0.99
1.52* *
* Significant at the 0.05 probability level. * * Significant at the 0.01 probability level. Abbreviations correspond with those given in Table 1. * 表示在0.05水平上显著相关; * * 表示在0.01水平上显著相关。
表2 云南和重庆321份甘蓝型油菜20个性状的方差分析 Table 2 Analysis of variance of 20 traits in 321B. napusaccessions cultivated at Chongqing and Yunnan
表3 云南和重庆321份甘蓝型油菜产量收获指数和产油量收获指数与17个产量组成性状的相关系数 Table 3 Correlation coefficients between YHI or OHI and 17 yield component traits in 321B. napusaccessions cultivated at Chongqing and Yunnan
性状 Trait
产量收获指数 YHI
产油量收获指数 OHI
云南Yunnan
重庆Chongqing
云南Yunnan
重庆Chongqing
单株平均千粒重 MTSWP
0.35* *
-0.01
0.37* *
-0.05
主序粒重 SWMI
0.27* *
0.69* *
0.16
0.42* *
一次粒重 SWPB
0.68* *
0.79* *
0.40* *
0.46* *
二次粒重 SWSB
0.40* *
0.46* *
0.39* *
0.24* *
单株角果数 SP
-0.08
0.45* *
0.01
0.31* *
主序角果数 MIS
-0.25* *
0.20*
-0.32* *
0.13
一次角果数 PBS
-0.16
0.52* *
-0.10
0.36* *
二次角果数 SBS
0.07
0.22*
0.16
0.11
主序千粒重 TSWMI
0.22*
0.08
0.28* *
-0.03
一次千粒重 TSWPB
0.30* *
0.10
0.34* *
0
二次千粒重 TSWSB
0.24* *
-0.15
0.33* *
-0.07
单株平均每角粒数 MSP
0.45* *
0.13
0.30* *
0.23*
主序每角粒数 SSMI
0.34* *
0.60* *
0.18*
0.41* *
一次每角粒数 SSPB
0.41* *
0.62* *
0.25* *
0.32* *
二次每角粒数 SSSB
0.19*
0.13
0.06
0.16
单株经济产量 PY
0.66* *
0.30* *
0.46* *
-0.02
单株生物产量 PB
-0.28* *
-0.52* *
-0.04
0.25* *
* Significant at the 0.05 probability level. * * Significant at the 0.01 probability level. Abbreviations correspond with those given in Table 1. * 表示在0.05水平上显著相关; * * 表示在0.01水平上显著相关。
表3 云南和重庆321份甘蓝型油菜产量收获指数和产油量收获指数与17个产量组成性状的相关系数 Table 3 Correlation coefficients between YHI or OHI and 17 yield component traits in 321B. napusaccessions cultivated at Chongqing and Yunnan
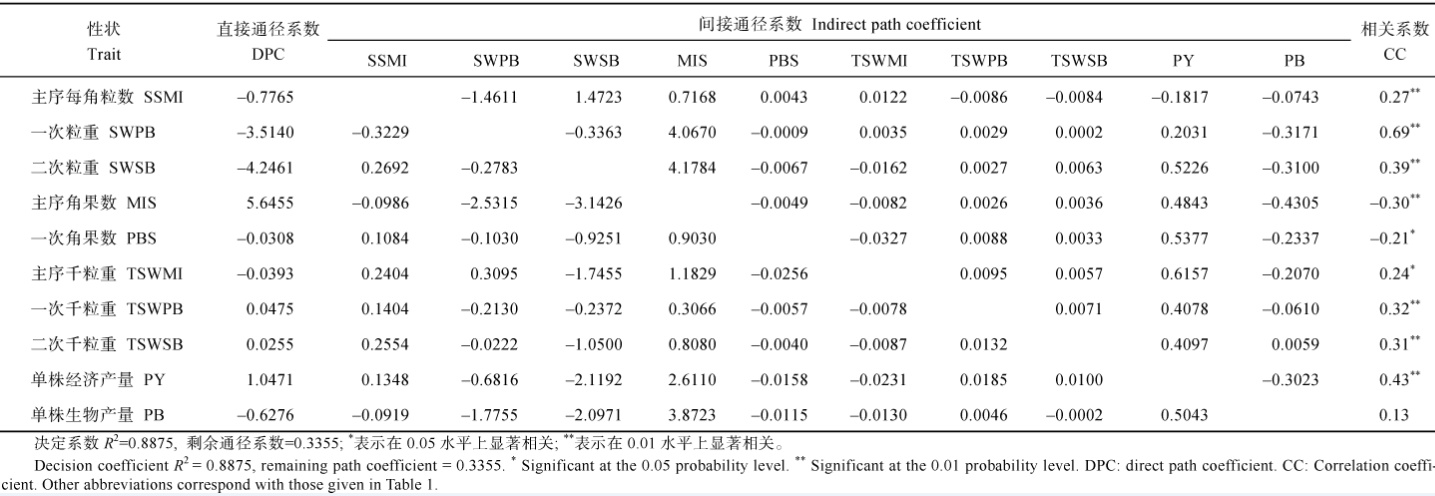
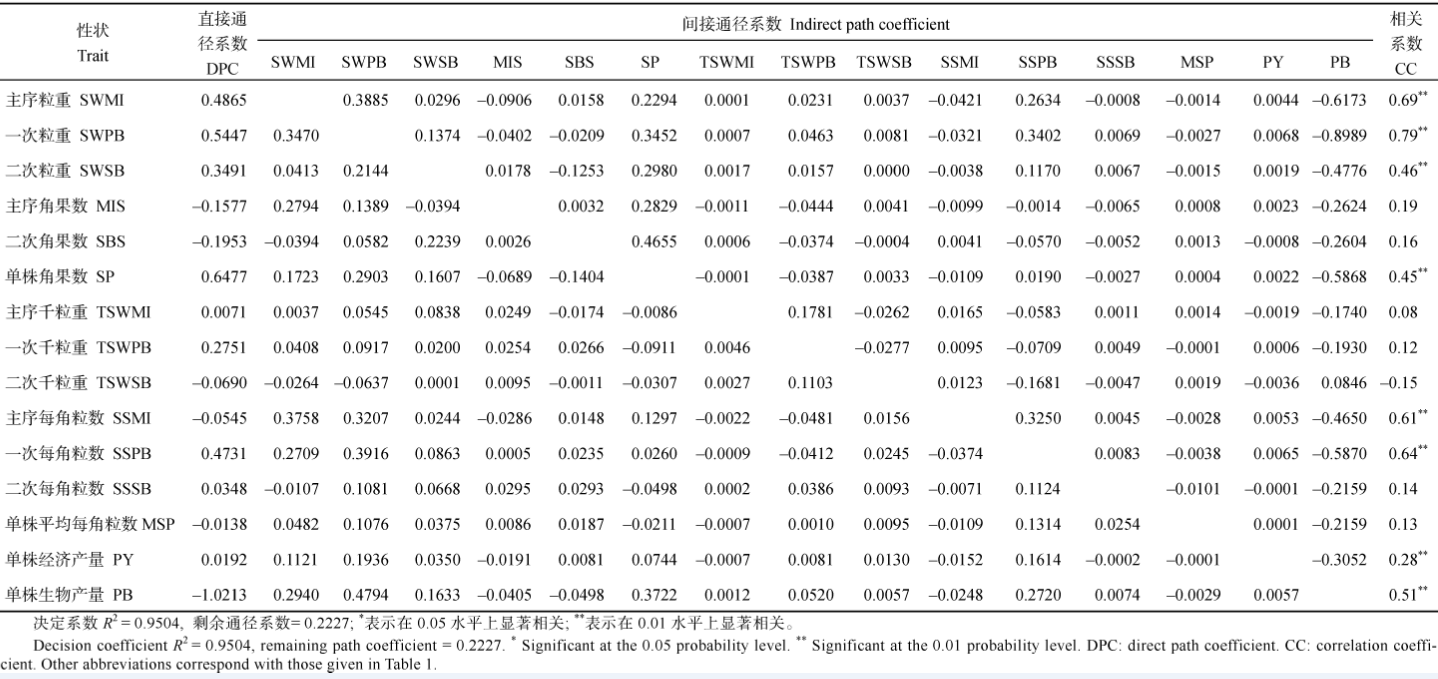
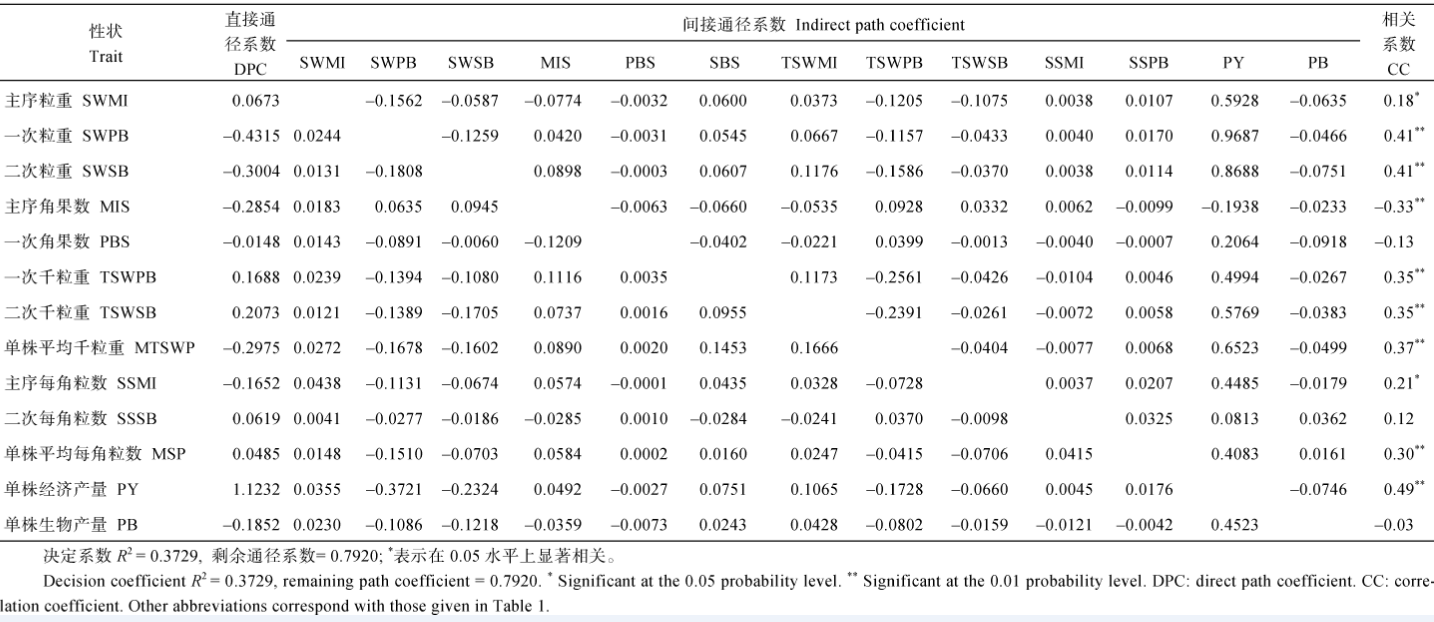
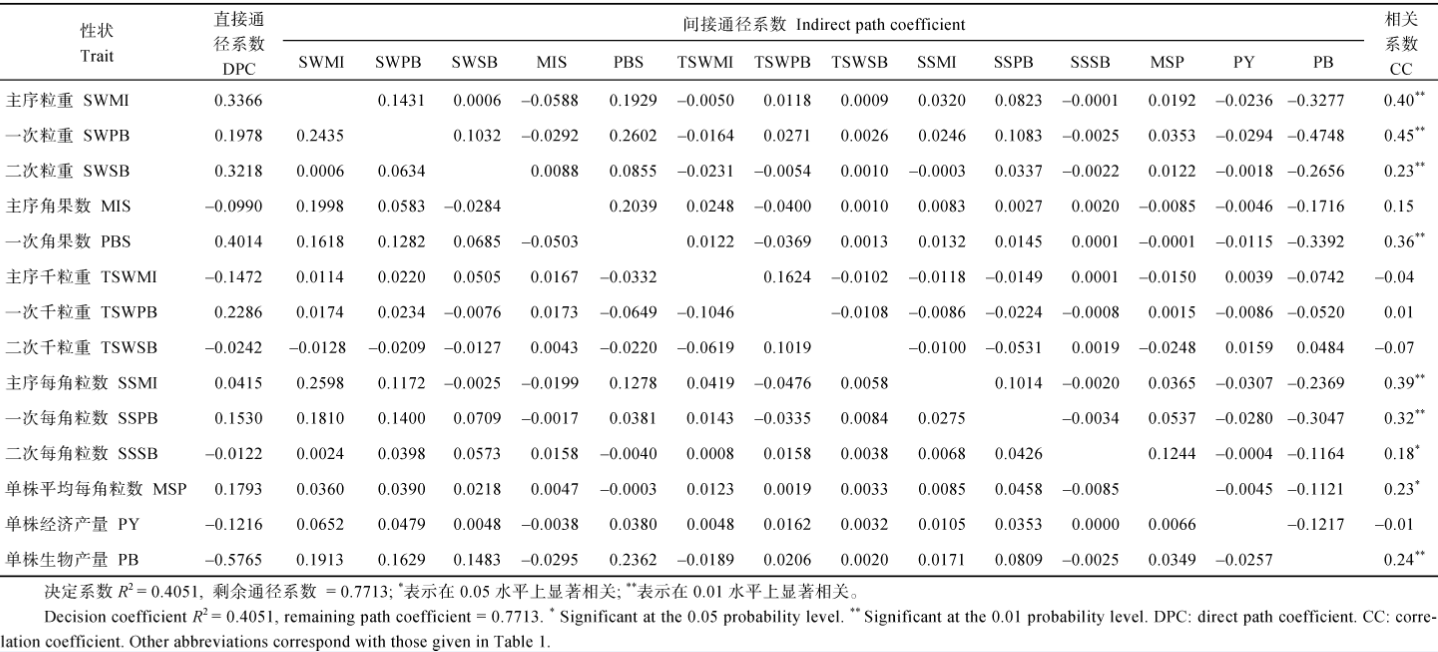
表7 重庆321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 7 Path analysis between OHI and 17 yield component traits in 321 B.napus accessions cultivated at ChongQing
表7 重庆321份甘蓝型油菜17个产量组成性状与产量收获指数的通径分析 Table 7 Path analysis between OHI and 17 yield component traits in 321 B.napus accessions cultivated at ChongQing
表8 Table 8 表8(Table 8)
表8 云南和重庆YHI或OHI差值与17个产量组成性状差值的相关分析 Table 8 Correlation coefficients between difference of YHI or OHI and difference of 17 yield component traits of Chongqing and Yunnan
性状 Trait
产量收获指数 YHI
产油量收获指数 OHI
性状 Trait
产量收获指数 YHI
产油量收获指数 OHI
主序粒重 SWMI
0.43* *
0.26* *
二次千粒重 TSWSB
0
0.07
一次粒重 SWPB
0.79* *
0.38* *
单株平均千粒重 MTSWP
0.10
0.24* *
二次粒重 SWSB
0.38* *
0.23* *
主序每角粒数 SSMI
0.48* *
0.26* *
主序角果数 MIS
-0.12
-0.20*
一次每角粒数 SSPB
0.45* *
0.23* *
一次角果数 PBS
0.19
0.02
二次每角粒数 SSSB
0.13
0.22*
二次角果数 SBS
0.10
-0.04
单株平均每角粒数 MSP
0.35* *
0.31* *
单株角果数 SP
0.17
-0.04
单株经济产量 PY
0.52* *
0.16
主序千粒重 TSWMI
0.08
0.28* *
单株生物产量 PB
0.15
0.08
一次千粒重 TSWPB
0.23*
0.32* *
* Significant at the 0.05 probability level. * * Significant at the 0.01 probability level. Abbreviations correspond with those given in Tables 1 and 4. * 表示在0.05水平上显著相关; * * 表示在0.01水平上显著相关。
表8 云南和重庆YHI或OHI差值与17个产量组成性状差值的相关分析 Table 8 Correlation coefficients between difference of YHI or OHI and difference of 17 yield component traits of Chongqing and Yunnan
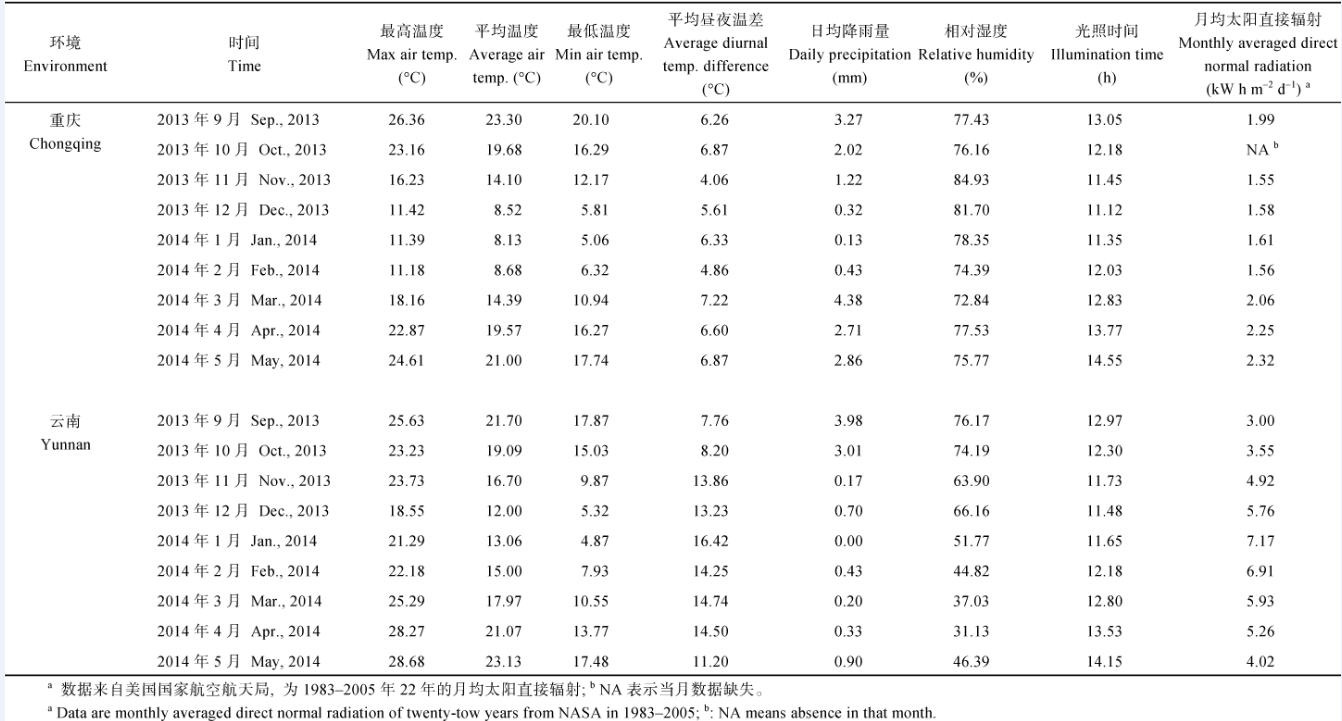
表9 Table 9 表9(Table 9)
表9 材料种植期间云南和重庆的气象资料 Table 9 Meteorological data at Chongqing and Yunnan during cultivation
表9 材料种植期间云南和重庆的气象资料 Table 9 Meteorological data at Chongqing and Yunnan during cultivation
蔡东芳. 甘蓝型油菜产量和品质相关性状关联分析. 华中农业大学博士学位论文, 湖北武汉, 2013. pp 1-2Cai DF. Association Mapping of Yield and Quality-related Traits in Brassica napus L. PhD Dissertation of Huazhong Agricultural University, Wuhan, China, 2013. pp 1-2 (in Chinese with English abstract)[本文引用:1]
[2]
李清波. 油菜不同品种及在不同种植密度及施氮条件下产量形成研究. 华中农业大学硕士学位论文, 湖北武汉, 2014. pp 2-8Li QB. Studies of the Yield Formation of Various Cultivars and under Different Planting Densities and Nitrogen Applications of Oilseed Rape. MS Thesis of Huazhong Agricultural University, Wuhan, China, 2014. pp 2-8 (in Chinese with English abstract)[本文引用:1]
[3]
Rand hirS, Malhotra SP, SinghR. Molecular manipulation of sourcesink interactions in crop plants. Proc Indian Natl Sci Acad, 1999, 6: 351-376[本文引用:1]
[4]
Reynolds MP, RajaramS, Sayre KD. Physiological and genetic changes of irrigated wheat in the post-green revolution period and approaches for meeting projected global demand . Crop Sci, 1999, 6: 1611-1621[本文引用:1]
[5]
Hay R KM. Harvest index: a review of its use in plant breeding and crop physiology. Annu Appl Biol, 1995, 126: 197-216[本文引用:2]
[6]
宋荷仙, 李跃建, 冯君成, 刘宗典, 杨永澄. 小麦收获指数和源、库性状的遗传研究. 中国农业科学, 1993, 26(3): 21-26Song HX, Li YJ, Feng JC, Liu ZD, Yang YC. Genetic research on harvest index and source sink traits of wheat. Sci Agric Sin, 1993, 26(3): 21-26 (in Chinese with English abstract)[本文引用:1]
[7]
何秀英, 陈钊明, 廖耀平, 程永盛, 陈粤汉. 水稻收获指数遗传及其与主要农艺性状的相关研究. 作物学报, 2006, 32: 911-916He XY, Chen ZM, Liao YP, Cheng YS, Chen YH. Heredity of rice harvest index and correlations between HI and main agronomic characters. Acta Agron Sin, 2006, 32: 911-916 (in Chinese with English abstract)[本文引用:1]
[8]
钟蕾. 不同收获指数型水稻品种产量构成整齐性及生育后期光合特性的差异性分析. 江西农业大学学报, 2012, 34: 627-634ZhongL. Comparative analysis on the yield component uniformity and photosynthesis characteristics during later growth stages in rice varieties with different harvest indexes. Acta Agric Univ Jiangxiensis (Nat Sci), 2012, 34: 627-634 (in Chinese with English abstract)[本文引用:1]
[9]
UnkovichM, BaldockJ, ForbesM. Variability in harvest index of grain crops and potential significance for carbon accounting: examples from Australian agriculture. Adv Agron, 2010, 105: 173-219[本文引用:1]
[10]
Kemanian AR, Stöckle CO, Huggins DR, Viega LM. A simple method to estimate harvest index in grain crops. Field Crops Res, 2007, 103: 208-216[本文引用:1]
[11]
黄露, 陶诗顺, 张敏, 姜磊, 彭雅利. 甘蓝型杂交油菜收获指数及其品种间差异性研究. 江苏农业科学, 2011, (1): 95-97HuangL, Tao SS, ZhangM, JiangL, Peng YL. Differences analysis of harvest and varieties of Brassica napus L. Jiangsu Agric Sci, 2011, (1): 95-97 (in Chinese with English abstract)[本文引用:1]
[12]
袁婺洲, 官春云. 影响油菜收获指数的几个生理因子. 作物学报, 1997, 23: 580-586Yuan WZ, Guan CY. Harvest index in rapeseed affected by a few physiological factors. Acta Agron Sin, 1997, 23: 580-586 (in Chinese with English abstract)[本文引用:1]
[13]
Yuan WZ, Guan CY. The variation of harvest index in Chinese oilseed rape. Life Sci Res, 1998, 3: 212-219[本文引用:2]
[14]
LuoX, MaC, YueY, HuK, LiY, DuanZ, WuM, TuJ, ShenJ, YiB, FuT. Unravelling the complex trait of harvest index in rapeseed (Brassica napus L. ) with association mapping. BMC Genomics, 2015, 16: 379[本文引用:1]
[15]
卢坤, 王腾岳, 徐新福, 唐章林, 曲存民, 贺斌梁颖, 李加纳. 甘蓝型油菜结角高度与荚层厚度的全基因组关联分析. 作物学报, 2016, 42: 344-352LuK, Wang TY, Xu XF, Tang ZL, Qu CM, HeB, LiangY, Li JN. Genome-wide association analysis of height of podding and thickness of pod canopy in Brassica napus. Acta Agron Sin, 2016, 42: 344-352 (in Chinese with English abstract)[本文引用:1]
[16]
Badger MR. Role of plant leaf development in optimising photosynthetic efficiency, capacity, growth and yield. In: Gready J E, Dwyer S A, Evans J R, eds. Applying Photosynthesis Research to Improvement of Food Crops. Canberra: Australian Centre for International Agricultural Research, 2009. pp 14-19[本文引用:1]
[17]
Evans JR. Improved photosynthetic efficiency is necessary to increase potential crop yield. In: Gready J E, Dwyer S A, Evans J R, eds. Applying Photosynthesis Research to Improvement of Food Crops. Canberra: Australian Centre for International Agricultural Research, 2009. pp 27-34[本文引用:1]
[18]
程永盛, 廖耀平, 何秀英, 陈钊明, 陈粤汉. 水稻收获指数与单株产量及其构成因素的相关和通径分析. 广东农业科学, 2006, (9): 36-38Cheng YS, Liao YP, He XY, Chen ZM, Chen YH. Path and coefficient analysis of harvest index, yield per plant and its components in rice. Guangdong Agric Sci, 2006, (9): 36-38 (in Chinese)[本文引用:2]
[19]
姚金保, 姚国才, 杨学明, 马鸿翔, 张平平. 小麦收获指数遗传及其与农艺性状的相关分析. 江苏农业学报, 2008, (1): 5-10Yao JB, Yao GC, Yang XM, Ma HX, Zhang PP. Inheritance of wheat harvest index and its correlations with agronomic traits. Jiangsu Agric Sci, 2008, (1): 5-10 (in Chinese with English abstract)[本文引用:1]
[20]
Asghari AR. Investigate the relationship and path coefficient analysis between yield and yield components of canola cultivars. Int J Farm Alli Sci, 2013, 2: 104[本文引用:2]
 , 申鸽子
, 申鸽子

{kind=link}