关键词:茶树; 越冬芽休眠; 物质交流; 钙黄素 Molecular Regulation and Substance Exchange Dynamics at Dormancy and Budbreak Stages in Overwintering Buds of Tea Plant TANG Hu1,2,**, HAO Xin-Yuan2,**, WANG Lu2, XIAO Bin1, WANG Xin-Chao2,*, YANG Ya-Jun1,2,* 1College of Horticulture, Northwest A&F University, Yangling 712100, China
2Tea Research Institute, Chinese Academy of Agricultural Sciences / National Center for Tea Improvement / Key Laboratory of Tea Plant Biology and Resources Utilization, Ministry of Agriculture, Hangzhou 310008, China
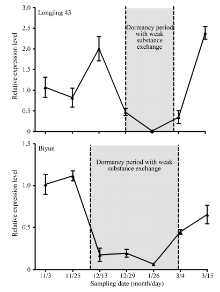
Fund:This study was supported by the National Natural Science Foundation of China (31370690), the China Agriculture Research System (CARS-23), and the Agricultural Science and Technology Innovation Program of CAAS (CAAS-ASTIP-2014-TRICAAS) AbstractEarly-sprouting cultivar Longjing 43 and late-sprouting cultivar Biyun were employed in this study to disclose the dormancy mechanism in tea plant with different sprouting phenophases. The levels of substance exchange were monitored by detecting the fluorescence signal in calcein treated overwintering buds. The glucanase related genes were identified by sequence homology analysis. Their characteristics and expression patterns during different time of winter were further analyzed. The substance exchanges were detected either in stem-bud unit or mother leaf-stem unit. From the initial formation to release in dormancy, the substance exchange in overwintering buds showed strong-weak-strong variation patterns in both cultivars, however, the duration of weak exchange stage was much shorter in Longjing 43 than in Biyun. Moreover, there was a close correlation between substance exchange variation pattern and the expression pattern of CsGLU1, a gene identified in tea plant with positive callose hydrolyzation activity. On the basis of promoter sequence analysis, plenty of transcription factor binding sequences related to hormone signaling, cold stimulation and dormancy regulation were found in CsGLU1 promoter region, which validates its putative functions in dormancy regulation. In conclusion, overwintering buds of tea plant have substance exchange with stem and mother leaf both in dormancy and non-dormancy status, furthermore, the variation of substance exchange level was consistent to the changes of dormancy status. CsGLU1 is a callose hydrolyzation related gene, which is supposed to be a key gene regulating tea plant dormancy transition through affecting the substance exchange in overwintering buds. The study provides meaningful results for understanding the changes of dormancy statuses in overwintering buds and deeply exploring the regulation mechanism in tea plant with different sprouting phenophase.
Keyword:Tea plant; Overwinter bud dormancy; Substances exchange; Calcein Show Figures Show Figures
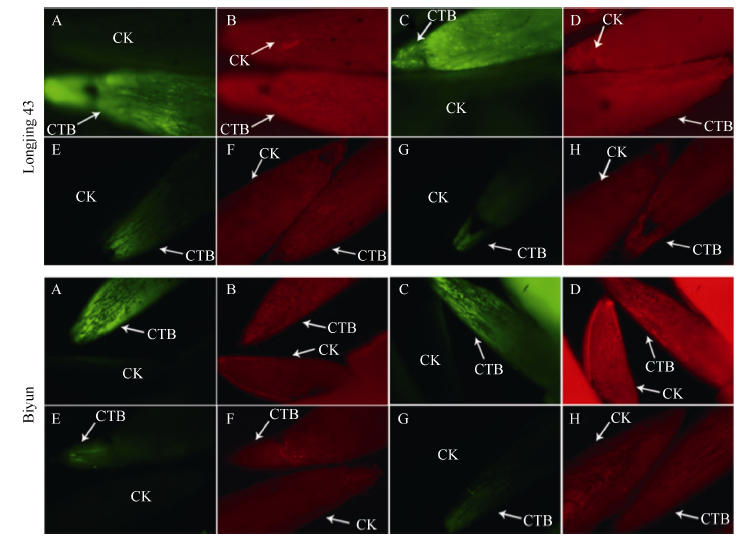
图1 龙井43及碧云越冬芽在休眠和活跃期的荧光观察“ 茎-芽” 处理越冬芽, A和B为活跃生长期(2015-10-31)样品, E和F为休眠期(2016-01-16)样品; “ 叶-芽” 处理越冬芽, C和D为活跃生长期(2015-10-31)样品, G和H为休眠期(2016-01-16)样品。4倍物镜观察, 绿色荧光为钙黄素在紫外光激发下的荧光信号, 红色荧光为越冬芽在绿光通道下的自发荧光。CTB: 钙黄素处理; CK: 水处理对照。Fig. 1 Fluorescence signals of overwintering buds at dormancy stage and active-growing stage in Longjing 43 and BiyunIn stem to bud treatment, samples A and B were active-growing buds (2015-10-31) and samples E and F were dormant buds (2016-01-16). In leaf to bud treatment, samples C and D were active-growing buds (2015-10-31) and samples G and H were dormant buds (2016-01-16). Pictures were taken under 4× objective lens. The green signals show fluorescence produced by calcein under UV light stimulation and the red signals show autofluorescence of bud under green light stimulation. CTB: calcein treatment; CK: control (H2O).
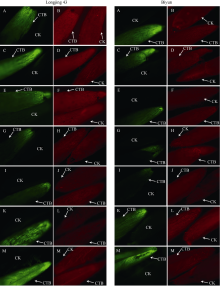
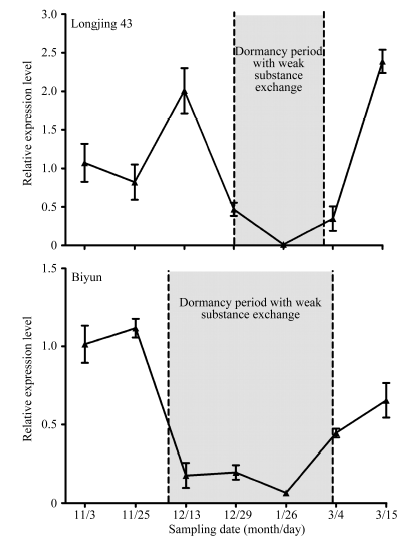
图2 冬季不同时期钙黄素处理(“ 茎-芽” 处理)的龙井43及碧云越冬芽荧光信号检测采样日期为2015-10-31 (A和B)、2015-11-16 (C和D)、2015-12-17 (E和F)、2016-01-06 (G和H)、2016-02-02 (I和J)、2016-02-25 (K和L)及2016-03-09 (M和N)。4倍物镜观察, 绿色荧光为钙黄素在紫外光激发下的荧光信号, 红色荧光为越冬芽在绿光通道下的自发荧光。CTB: 钙黄素处理; CK: 水处理对照。Fig. 2 Fluorescence signals of calcein treated overwintering buds (stem to bud treatment) of Longjing 43 and Biyun during different periods of winterThe sampling dates were 2015-10-31 (A and B), 2015-11-16 (C and D), 2015-12-17 (E and F), 2016-01-06 (G and H), 2016-02-02 (I and J), 2016-02-25 (K and L), and 2016-03-09 (M and N). Pictures were taken under 4× objective lens. The green signals show fluorescence produced by calcein under UV light stimulation and the red signals show autofluorescence of bud under green light stimulation. CTB: calcein treatment; CK: control (H2O).
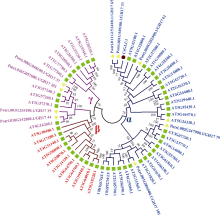
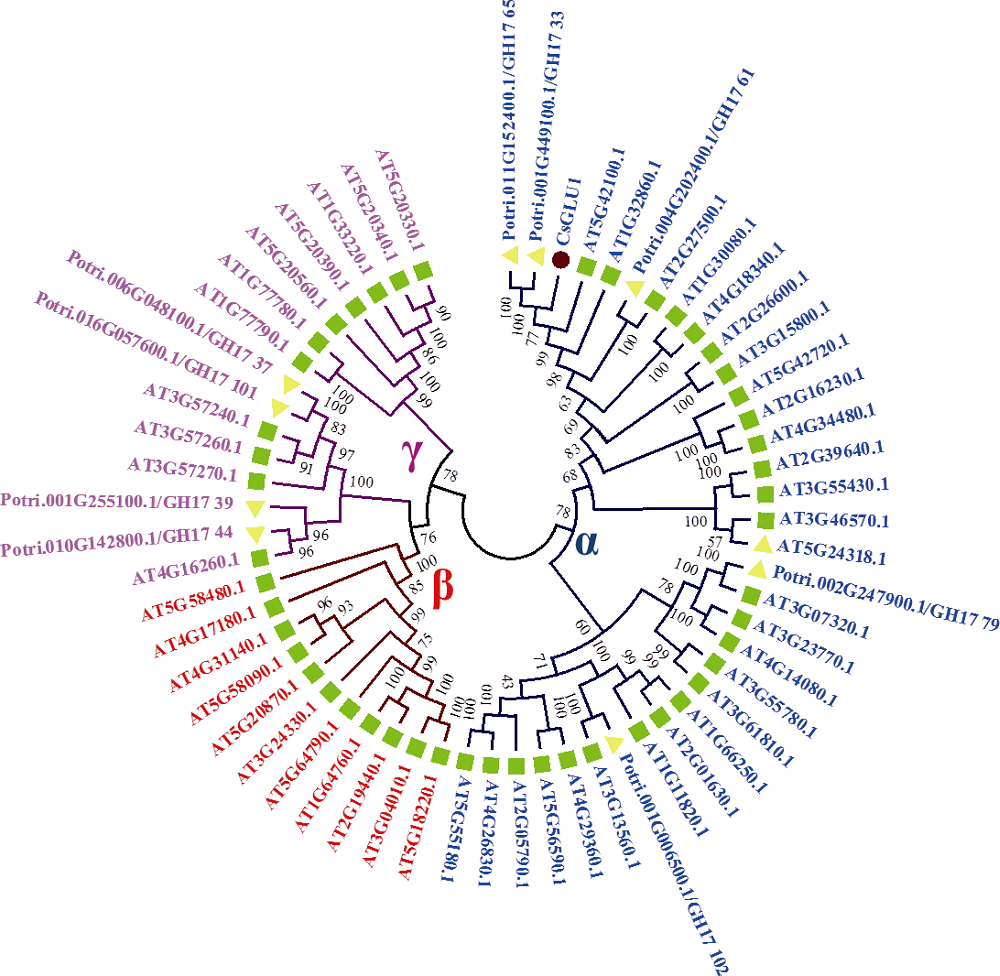
图3 CsGLU1与拟南芥和杨树葡聚糖酶家族成员的系统发育树分析蓝色、红色和紫色字体分别显示α 、β 和γ 亚家族基因。■拟南芥葡聚糖酶基因家族; ▼杨树葡聚糖酶基因家族; ●茶树CsGUL1。Fig. 3 Phylogenetic tree analysis of CsGUL1 with glucanase family members from Arabidopsis and Populusα -, β -, and γ -clade genes are in blue, red, and purple font, respectively. ■ Genes from Arabidopsis; ▼ Genes from Populus; ● CsGLU1.
RuttinkT, ArendM, MorreelK, StormeV, Rombaut sS, Fromm J, Bhalerao R, Boerjan W, Rohde A. A molecular timetable for apical bud formation and dormancy induction in poplar. Plant Cell, 2007, 19: 2370-2390[本文引用:2]
[2]
Cooke JE, Eriksson ME, JunttilaO. The dynamic nature of bud dormancy in trees: environmental control and molecular mechanisms. Plant Cell Environ, 2012, 35: 1707-1728[本文引用:3]
[3]
Rinne PL, van der SchootC. Symplasmic fields in the tunica of the shoot apical meristem coordinate morphogenetic events. Development, 1998. 125: 1477-1485[本文引用:1]
[4]
NathalieD, AnnikaJ, BabaK, SchraderJ, SjödinA, Bhalerao RR, ResmanL, TryggJ, MoritzT, Bhalerao RP. Environmental and hormonal regulation of the activity-dormancy cycle in the cambial meristem involves stage-specific modulation of transcriptional and metabolic networks. Plant J, 2007, 50: 557-573[本文引用:1]
[5]
van der SchootC, Rinne PL. Dormancy cycling at the shoot apical meristem: transitioning between self-organization and self-arrest. Plant Sci, 2011, 180: 120-131[本文引用:1]
[6]
Low HP, GrécoB, TanahashiY, GallantJ, Jones SN, Billings- GagliardiS, Recht LD, Schwartz WJ. Plasmodesmata at the crossroads between development, dormancy, and defense. Can J Bot, 2004, 81: 1182-1197[本文引用:1]
[7]
Rinne PL, WellingA, VahalaJ, RipelL, RuonalaR, Kangasjarvi J, van der Schoot C. Chilling of dormant buds hyperinduces FLOWERING LOCUS T and recruits GA-inducible 1, 3-beta-glucanases to reopen signal conduits and release dormancy in Populus. Plant Cell, 2011, 23: 130-146[本文引用:3]
[8]
Rinne PL, Kaikuranta P M, van der Schoot C. The shoot apical meristem restores its symplasmic organization during chilling- induced release from dormancy. Plant J, 2001, 26: 249-264[本文引用:2]
[9]
晏嫦妤, 李家贤, 黄华林, 何雨媚. 茶树休眠的研究进展. 安徽农业科学, 2012, 40: 10387-10389Yan CY, Li JX, Huang HL, He YM. Research progress of tea plant dormancy. J Anhui Agric Sci, 2012, 40: 10387-10389 (in Chinese with English abstract)[本文引用:1]
[10]
钱利生, 沈生荣, 潘根生. 茶树新梢内源激素的HPLC分析及日变化. 茶叶科学, 1996, (2): 135-139Qian LS, Shen SR, Pan GS. HPLC analysis and diurnal variation of the endogenous hormones in shoots of tea plants. J Tea Sci, 1996, (2): 135-139 (in Chinese with English abstract)[本文引用:1]
[11]
黄亚辉, 粟本文, 郑红发, 曾贞, 刘霞林. 茶树春梢萌动期间内源激素含量的变化. 植物生理学通讯, 2001, 37: 306-307Huang YH, Su BW, Zheng HF, ZengZ, Liu XL. Changes of endohormone in prouting shoot of tea plant. Plant Physiol Commun, 2001, 37: 306-307 (in Chinese with English abstract)[本文引用:1]
[12]
禹利君, 史云峰, 肖海云, 刘富知, 刘仲华. 不同物候型茶树内源GA3和ABA的变化及其对腋芽萌发调控的影响. 作物学报, 2008, 34: 277-283Yu LJ, Shi YF, Xiao HY, Liu FZ, Liu ZH. Dynamic changes of endogenous GA3 and ABA contents in tea culticars with different phenological characters and their impact on the regulation axillary buds sprouting. Acta Agron Sin, 2008, 34: 277-283 (in Chinese with English abstract)[本文引用:1]
[13]
WangX, HaoX, MaC, CaoH, YueC, WangL, ZengJ, YangY. Identification of differential gene expression profiles between winter dormant and sprouting axillary buds in tea plant (Camellia sinensis) by suppression subtractive hybridization. Tree Genet Genom, 2014, 10: 1149-1159[本文引用:1]
[14]
王新超, 杨亚军, 马春雷, 金基强, 曹红利. 茶树细胞周期蛋白基因的克隆与表达. 西北植物学报, 2011, 31: 2365-2372Wang XC, Yang YJ, Ma CL, Jin JQ, Cao HL. Cloning and expression analysis of cyclin gene (CsCYC1) of tea plant. Acta Bot Boreali-Occident Sin, 2011, 31: 2365-2372 (in Chinese with English abstract)[本文引用:1]
[15]
王新超, 马春雷, 杨亚军, 金基强, 马建强, 曹红利. 茶树细胞周期蛋白依赖激酶(CsCDK)基因cDNA全长克隆与分析. 园艺学报, 2012, 39: 333-342Wang XC, Ma CL, Yang YJ, Jin JQ, Ma JQ, Cao H L. cDNA cloning and expression analysis of cyclin-dependent kinase (CsCDK) gene in tea plant. Acta Hort Sin, 2012, 39: 333-342 (in Chinese with English abstract)[本文引用:1]
[16]
Doxey AC, Yaish MW, Moffatt BA, GriffithM, McConkey B J. Functional divergence in the Arabidopsis beta-1, 3-glucanase gene family inferred by phylogenetic reconstruction of expression states. Mol Biol Evol, 2007, 24: 1045-1055[本文引用:2]
[17]
Rinne PL, Paul LK, VahalaJ, Kangasjarvi J, van der Schoot C. Axillary buds are dwarfed shoots that tightly regulate GA pathway and GA-inducible 1, 3-beta-glucanase genes during branching in hybrid aspen. J Exp Bot, 2016, 67: 5975-5991[本文引用:4]
[18]
ChangS, PuryearJ, CairneyJ. A simple and efficient method for isolating RNA from pine trees. Plant Mol Biol Rep, 1993, 11: 113-116[本文引用:1]
[19]
HaoX, Horvath DP, Chao WS, YangY, WangX, XiaoB. Identification and evaluation of reliable reference genes for quantitative real-time PCR analysis in tea plant (Camellia sinensis (L. ) O. Kuntze). Int J Mol Sci, 2014, 15: 22155-22172[本文引用:1]
[20]
Jian LC, Li PH, Sun LH, Chen T H H. Alterations in ultrastructure and subcellular localization of Ca2+ in poplar apical bud cells during the induction of dormancy. J Exp Bot, 1997, 48: 1195-1207[本文引用:1]
[21]
刘芳, 王家艳, 王晓丽, 周蕴薇. 细叶百合鳞茎在低温解除休眠过程中茎尖细胞超微结构的变化. 园艺学报, 2013, 40: 1110-1118LiuF, Wang JY, Wang XL, Zhou YW. The apical bud cell ultra-structure changes of Lilium pumilum bulbs during breaking dormancy under refrigerated condition. Acta Hort Sin, 2013, 40: 1110-1118 (in Chinese with English abstract)[本文引用:1]
[22]
RuonalaR, Rinne PL, Kangasjärvi J, van der Schoot C. CENL1 expression in the rib meristem affects stem elongation and the transition to dormancy in Populus. Plant Cell, 2008, 20: 59-74[本文引用:1]
[23]
Knox JP, Benitez-AlfonsoY. Roles and regulation of plant cell walls surrounding plasmodesmata. Curr Opin Plant Biol, 2014, 22: 93-100[本文引用:1]
[24]
Paul LK, Rinne P L, van der Schoot C. Refurbishing the plasmodesmal chamber: a role for lipid bodies?Front Plant Sci, 2014. 5: 40[本文引用:1]
[25]
Nagar PK, KumarA. Changes in endogenous gibberellin activity during winter dormancy in tea (Camellia sinensis (L. ) O. Kuntze). Acta Physiol Plant, 2000, 22: 439-443[本文引用:1]
[26]
CaoH, WangL, YueC, HaoX, WangX, YangY. Isolation and expression analysis of 18 CsbZIP genes implicated in abiotic stress responses in the tea plant (Camellia sinensis). Plant Physiol Biochem, 2015, 97: 432-442[本文引用:1]
[27]
Wang XC, Zhao QY, Ma CL, Zhang ZH, Cao HL, Kong YM, YueC, Hao XY, ChenL, Ma JQ, Jin JQ, LiX, Yang YJ. Global transcriptome profiles of Camellia sinensis during cold acclimation. BMC Genom, 2013, 14: 415[本文引用:1]
[28]
郝心愿, 曹红利, 杨亚军, 王新超, 马春雷, 肖斌. 茶树生长素响应因子基因CsARF1的克隆与表达分析. 作物学报, 2013, 39: 389-397Hao HX, Cao CH, Yang YJ, Wang XC, Ma CL, XiaoB. Cloning and expression analysis of auxin response factor gene (CsARF1) in tea plant (Camellia sinensis [L. ] O. Kuntze). Acta Agron Sin, 2013, 39: 389-397 (in Chinese with English abstract)[本文引用:1]
[29]
JimenezS, Lawton-Rauh A L, Reighard G L, Abbott A G, Bielenberg D G. Phylogenetic analysis and molecular evolution of the dormancy associated MADS-box genes from peach. BMC Plant Biol, 2009, 9: 81[本文引用:1]
 , 郝心愿
, 郝心愿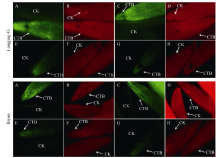


{kind=link}
{kind=link}
{kind=link}
{kind=link}