关键词:水稻; 淡黄叶矮化突变体; 遗传分析; 基因定位 Genetic Analysis and Gene Mapping of Yellow leaf and dwarf (yld) Mutant in Rice LI Zi-Zhuang1,2,**, XU Qian-Kun2,**, YU Hai-Ping2, ZHOU Ting-Ting2, XUE Da-Wei1, ZENG Da-Li2, GUO Long-Biao2, QIAN Qian2,*, REN De-Yong2,* 1College of Life and Environmental Sciences, Hangzhou Normal University, Hangzhou 310006, China
2State Key Laboratory of Rice Biology, China National Rice Research Institute, Hangzhou 310006, China
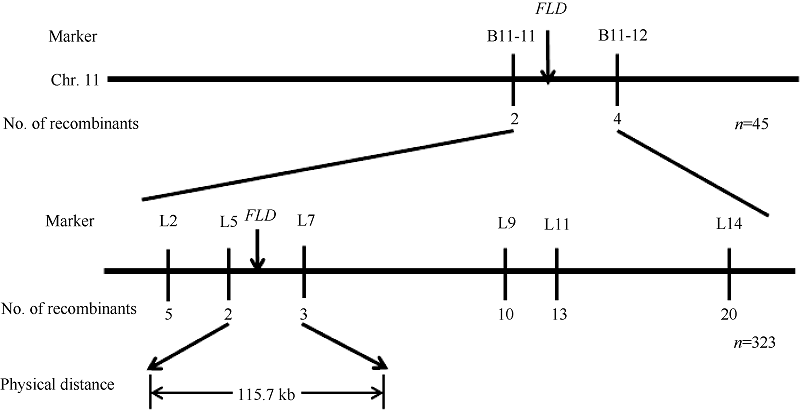
Fund:The study was supported by the National Natural Science Foundation of China (31401464). AbstractLeaf color mutants of rice are ideal materials in studies on photosynthesis, chlorophyll metabolism and chloroplast development in plants. A yellow leaf and dwarf mutant, Yellow leaf and dwarf( yld), was obtained from ethyl methane sulfonate (EMS)-treated Shuhui 527 ( Oryza sativa L.). Compared with the wild type, the yld mutant showed yellow leaf and dwarfism, and the contents of chlorophyll and carotenoid were obviously decreased. Transmission electron microscope observation revealed that the structure of most chloroplasts seemed to be normal, however, with the fuzzy grana, and fewer and looser stroma lamella in the yld mutant. Meantime, plant height, branch number, grain number per panicle, 1000-grain weight and seed-setting rate were significantly decreased, while the number of effective panicle was obviously increased in the yld mutant compared with those in the wild type. Genetic analysis showed that the yld mutant traits were controlled by a single recessive gene. The 323 mutational individuals from the F2 generation of the cross of Wuyunjing 7 and yld mutant were used for gene mapping. Finally, the YLDlocus was mapped on chromosome 11 between two Indel markers L5 and L7, with an approximate 115 kb physical region. These results would facilitate cloning and functional analysis for the YLD gene.
Keyword:Rice; Yellow leaf and dwarf mutant; Genetic analysis; Gene mapping Show Figures Show Figures
图1 野生型(左)和yld突变体(右)表型 A: 苗期; B: 分蘖期; C: 抽穗期; D: 剑叶; E: 倒二叶; F: 倒三叶。Fig. 1 Phenotypes of the wild type (left) and yld mutant (right) A: seedling stage; B: tillering stage; C: heading stage; D: flag leaf; E: second leaf from top; F: third leaf from top. Bar=10 cm.
表2 Table 2 表2(Table 2)
表2 野生型和yld突变体农艺性状 Table 2 Agronomic traits of the wild type (WT) and theyld mutant
材料 Material
株高 Plant height (cm)
有效穗数 Effective panicle
穗粒数 Grain number per panicle
结实率 Seed-setting rate (%)
千粒重 1000-grain weight (g)
一次枝梗 Primary branch number
二次枝梗 Secondary branch number
WT
104.4± 4.12
13.0± 2.51
203.0± 11.00
89.3± 5.55
33.9± 0.17
10.9± 1.00
33.9± 6.00
yld
64.6± 4.50* *
25.3± 3.51* *
52.9± 6.13* *
76.9± 3.90* *
28.6± 0.16*
7.8± 0.50* *
5.4± 0.50
* Represents significant difference between the yldmutant and wild type at the 0.05 probability level. * * Represents significant difference between the yldmutant and wild type at the 0.01 probability level. * 表示野生型与突变体在0.05水平差异显著; * * 表示野生型与突变体在0.01水平差异显著。
表2 野生型和yld突变体农艺性状 Table 2 Agronomic traits of the wild type (WT) and theyld mutant
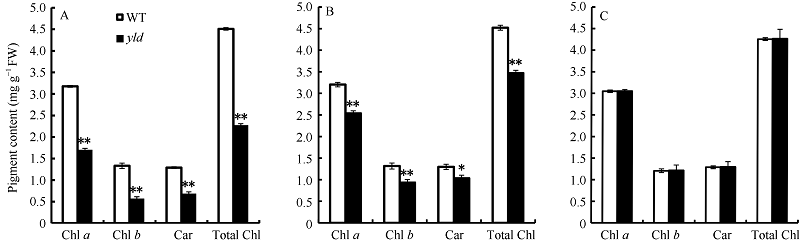
图2 野生型和yld突变体的剑叶、倒二叶及倒三叶光合色素含量比较 A: 剑叶光合色素含量比较; B: 倒二叶光合色素含量比较; C: 倒三叶光合色素含量比较。Chl a: 叶绿素a; Chl b: 叶绿素b; Total Chl: 叶绿素a+叶绿素b; Car: 类胡萝卜素。* 表示野生型与突变体在0.05水平差异显著; * * 表示野生型与突变体在0.01水平差异显著。Fig. 2 Comparison of photosynthetic pigments of the flag, second and third leaf in the yld mutant and wild type A: comparison of photosynthetic pigments of the flag leaf; B: comparison of photosynthetic pigments of the second leaf; C: comparison of photosynthetic pigments of the third leaf. Chl a: chlorophyll a; Chl b: chlorophyll b; Total Chl: content of chlorophyll aand chlorophyll b; Car: carotenoids. * represents significant difference between theyldmutant and wild type at the 0.05 probability level; * * represents significant difference between theyldmutant and wild type at the 0.01 probability level.
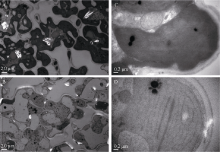
图3 野生型和yld突变体的细胞超微结构 A: 野生型叶肉细胞; B: yld突变体叶肉细胞; C: 野生型叶绿体; D: yld突变体叶绿体。Fig. 3 Cell ultrastructure of the wild type and yldmutant A: mesophyll cell of the wild type; B: mesophyll cell of theyldmutant; C: chloroplast of the wild type; D: chloroplast of the yld mutant.
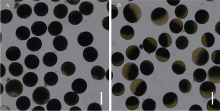
图4 野生型和yld突变体花粉育性 A: 野生型花粉; B: yld突变体花粉。Fig. 4 Pollen fertility in the wild type and yld mutant A: pollen fertility in the wild type; B: pollen fertility in the yld mutant. Bar = 50 μ m.
MoritaR, SatoY, MasudaY, NishimuraM, KusabaM. Defect innon-yellow coloring 3, an a/b hydrolase-fold family protein, causes a stay-green phenotype during leaf senescence in rice. Plant J, 2009, 59: 940-952[本文引用:1]
[2]
WuZ, ZhangX, HeB, DiaoL, ShengS, WangJ, GuoX, SuN, WangL, JiangL, WangC, ZhaiH, WanJ. A chlorophyll- deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
[3]
Larkin RM, Alonso JM, Ecker JR, ChoryJ. GUN4, a regulator of chlorophyll synthesis and intracellular signaling. Science, 2003, 299: 902-906[本文引用:1]
[4]
SugimotoH, KusumiK, TozawaY, YazakiJ, KishimotoN, KikuchiS, IbaK. The virescent-2 mutation inhibits translation of plastid transcripts for the plastidgenetic system at an early stage of chloroplast differentiation. Plant Cell Physiol, 2004, 45: 985-996[本文引用:1]
[5]
KusumiK, SakataC, NakamuraT, KawasakiS, YoshimuraA, IbaK. A plastid protein NUS1 is essential for build-up of the genetic system for early chloroplast development under cold stress conditions. Plant J, 2011, 68: 1039-1050[本文引用:1]
[6]
Goh CH, SatohK, KikuchiS, Kim SC, Ko SM, Kang HG, Jeon JS, Kim CS, Park YI. Mitochondrial activity in illuminated leaves of chlorophyll-deficient mutant rice (OsCHLH) seedlings. Plant Biotechnol Rep, 2010, 4: 281-291[本文引用:1]
[7]
Awan MA, Konzak CF, Rutger JN, Nilan RA. Mutagenic effects of sodium azide in rice. Crop Sci, 1979, 20: 663-668[本文引用:1]
[8]
LeeS, Kim JH, Yoo ES, Lee CH, HirochikaH, AnG. Differential regulation of chlorophyll a oxygenase genes in rice. Plant Mol Biol, 2005, 57: 805-818[本文引用:1]
[9]
刘文真. 三个水稻叶色突变体的鉴定与基因克隆. 浙江大学博士学位论文, 浙江杭州, 2006Liu WZ. Characterization of Three Chlorophyll Deficient Mutants in Rice. PhD Dissertation of Zhejiang University, Hangzhou, China, 2006 (in Chinese with English abstract)[本文引用:1]
[10]
Wu ZM, ZhangX, HeB, Diao LP, Sheng SL, Wang JL, Guo XP, SuN, Wang LF, JiangL, Wang CM, Zhai HQ, Wan JM. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 145: 29-40[本文引用:1]
[11]
Jung KH, HurJ, Ryu CH, ChoiY, Chung YY, MiyaoA, HirochikaH, AnG. Characterization of a rice chlorophyll-deficient mutant using the T-DNA gene-trap system. Plant Cell Physiol, 2003, 44: 463-472[本文引用:1]
[12]
Zhang HT, Li JJ, Yoo JH, Yoo SC, Cho SH, Koh HJ, Seo HS, Paek NC. Rice Chlorina-1 and Chlorina-9 encode Chl D and Chl I subunits of Mg-chelatase, a key enzyme for chlorophyll synthesis and chloroplast development. Plant Mol Biol, 2006, 62: 325-337[本文引用:1]
[13]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, KawasakiS, TanakaR, HirochikaH, NishimuraM, TanakaA. Rice Non-yellow Coloring 1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1]
[14]
SatoY, MoritaR, KatsumaS, NishimuraM, TanakaA, KusabaM. Two short-chain dehydrogenase/reductases, Non-yellow Coloring 1 and Nyc1-like, are required for chlorophyll b and light-harvesting complex II degradation during senescence in rice. Plant J, 2009, 57: 120-131[本文引用:1]
[15]
Zhang ZM, Tan JJ, Shi ZY, Xie QJ, XingY, Liu CH, Chen QL, Zhu HT, WangJ, Zhang JL, Zhang GQ. Albino leaf 1 that encodes the sole octotricopeptide repeat protein is responsible for chloroplast development in rice. Plant Physiol, 2016, 171: 1182-1191[本文引用:1]
[16]
Li CM, HuY, HuangR, Ma XZ, WangY, Liao TT, ZhongP, Xiao FL, Sun CH, Xu ZJ, Deng XJ, Wang PR. Mutation of FdC2 gene encoding a ferredoxin-like protein with C-terminal extension causes yellow-green leaf phenotype in rice. Plant Sci, 2015, 238: 127-134[本文引用:1]
[17]
Yang YL, XuJ, Huang LC, Leng YJ, Dai LP, Rao YC, ChenL, Wang YQ, Tu ZJ, HuJ, Ren DY, Zhang GH, ZhuL, Guo LB, QianQ, Zeng DL. PGL, encoding chlorophyllide a oxygenase 1, impacts leaf senescence and indirectly affects grain yield and quality in rice. J Exp Bot, 2015, 67: 1297-1310[本文引用:2]
[18]
SakurabaY, Rahman ML, Cho SH, Kim YS, Koh HJ, Yoo SC, Paek NC. The rice faded green leaf locus encodes protochlorophyllide oxidoreductase B and is essential for chlorophyll synthesis under high light conditions. Plant J, 2013, 74: 122-133[本文引用:1]
[19]
FangJ, Chai CL, QianQ, Li CL, Tang JY, SunL, Huang ZJ, Guo XL, Sun CH, LiuM, ZhangY, Lu QT, Wang YQ, Lu CM, HanB, ChenF, Cheng ZK, Chu CC. Mutations of genes in synthesis of the carotenoid precursors of ABA lead to pre-harvest sprouting and photo-oxidation in rice. Plant J, 2008, 54: 177-189[本文引用:1]
[20]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomenbranes. Meth Enzymol, 1987, 148: 350-382[本文引用:1]
[21]
何瑞峰, 丁毅, 余金洪, 祖明生. 水稻温敏叶绿素突变体叶片超微结构的研究. 武汉植物学研究, 2001, 19: 1-5He RF, DingY, Yu JH, Zu MS. Study on leaf ultrastructure of the thermo-sensitive chlorophyll deficient mutant in rice. J Wuhan Bot Res, 2001, 19: 1-5 (in Chinese with English abstract)[本文引用:1]
[22]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1]
[23]
Rogers SO, Bendich AJ. Extraction of DNA from milligram amounts of fresh, herbarium and mummified plant tissues. Plant Mol Biol, 1985, 5: 69-76[本文引用:1]
[24]
Kosambi DD. The estimation of map distances from recombination values. Ann Hum Genet, 1944, 12: 172-175[本文引用:1]
[25]
王亚琴, 施军琼, 张婷, 李燕, 张天泉, 张小龙, 桑贤春, 凌英华, 何光华. 水稻绿叶突变体ygl13的鉴定及候选基因分析. 中国农业科学, 2015, 48: 4197-4208Wang YQ, Shi JQ, ZhangT, LiY, Zhang TQ, Zhang XL, Sang XC, Ling YH, He GH. Characterization and cand idate gene analysis of yellow-green leaf mutant ygl13 in rice. Sci Agric Sin, 2015, 48: 4197-4208 (in Chinese with English abstract)[本文引用:1]
[26]
Zhang FT, Luo XD, Hu BL, WanY, Xie JK. YGL138(t), encoding a putative signal recognition particle 54 kDa protein, is involved in chloroplast development of rice. Rice, 2013, 6: 7[本文引用:1]
[27]
Mao DH, Yu HH, Liu TM, Yang GY, Xing YZ. Two complementary recessive genes in duplicated segments control etiolation in rice. Theor Appl Genet, 2011, 122: 373-383[本文引用:1]
[28]
Han SH, SakurabaY, Koh HJ, Paek NC. Leaf variegation in the rice zebra2 mutant is caused by photoperiodic accumulation of tetra-cis-lycopene and singlet oxygen. Mol Cells, 2012, 33: 87-97[本文引用:2]
[29]
邢才, 王贵学, 黄俊丽, 吴金钟. 植物叶绿素突变体及其分子机理的研究进展. 生物技术通报, 2008, (5): 10-12XingC, Wang GX, Huang JL, Wu JZ. Research on chlorophyll mutation of plants and molecular mechanism. Biotechnol Bull, 2008, (5): 10-12 (in Chinese with English abstract)[本文引用:1]
 , 徐乾坤
, 徐乾坤






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}