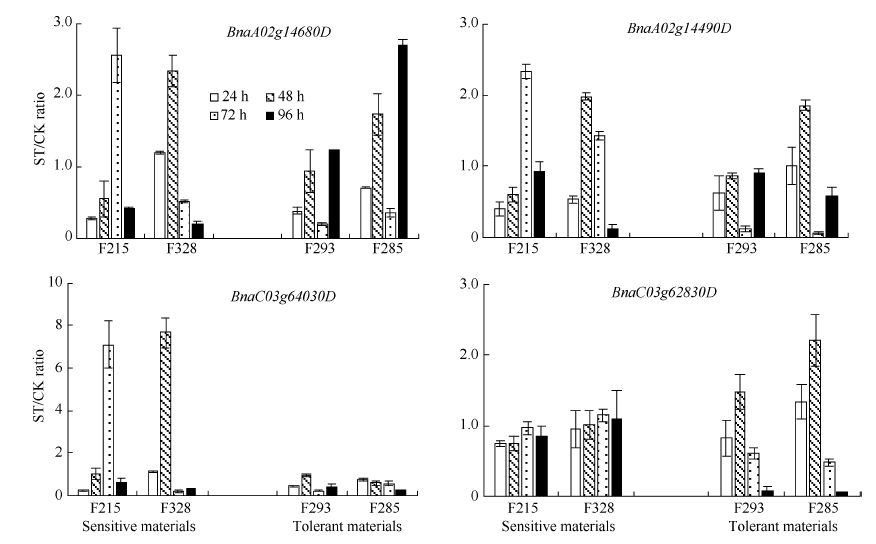
关键词:甘蓝型油菜; 盐胁迫; 数量性状位点; 单核苷酸多态性; 候选基因 QTL Mapping for Seedling Dry Weight and Fresh Weight under Salt Stress and Candidate Genes Analysis in Brassica napusL HOU Lin-Tao**, WANG Teng-Yue**, JIAN Hong-Ju, WANG Jia, LI Jia-Na, LIU Lie-Zhao* College of Agronomy and Biotechnology, Southwest University, Chongqing 400715, China Fund:The study was supported by the National Natural Science Foundation of China (31371655) AbstractSalt stress is one of the main abiotic stresses affecting crop yield and it would be very important by using the salt tolerance related markers in rapeseed breeding to improve the oilseed production. In this research, the Brassica napus L. high generation recombinant inbred lines (RIL) population derived from the cross of GH06 and P174 via single seed descent propagation was used for QTL mapping and candidate gene analysis. The fresh and dry weight of leaf and root were measured at 25 days after the seedlings were grown in Hoagland solution with 16 g L-1NaCl. Composite interval mapping (CIM) was used to identify the related QTLs according to the high density SNP genetic map, and the candidate gene expression in the extreme lines tested by qRT-PCR. A total of 19 QTLs were identified in the control and salt stress treatment, and six QTLs were mapped on chromosomes A02, A04, and C03 under salt stress, with contribution rate ranged from 7.16% to 16.15%. Eight genes were identified according to the BLAST of genes in the QTL confidence intervals and the salt stress related genes in Arabidopsis. The expression of four candidate genes in the extreme lines showed that BnaA02g14680D and BnaA02g14490D under salt stress treatment for 48 or 72 hours had higher expression than the control, which indicates that the expressions are induced by salt stress. The relative expressions of gene BnaC03g64030Din sensitive extreme lines were higher than those in tolerant extreme lines. There were no changed in expression for gene BnaC03g62830D in sensitive extreme lines but increased expression at 48 hours and reduced expression at 72 hours after salt treatment in tolerant extreme lines, showing the enhance of plant salt tolerance possibly. Our research laid a foundation for the function research of salt tolerant gene in rapeseed and the breeding of salt tolerant rapeseed.
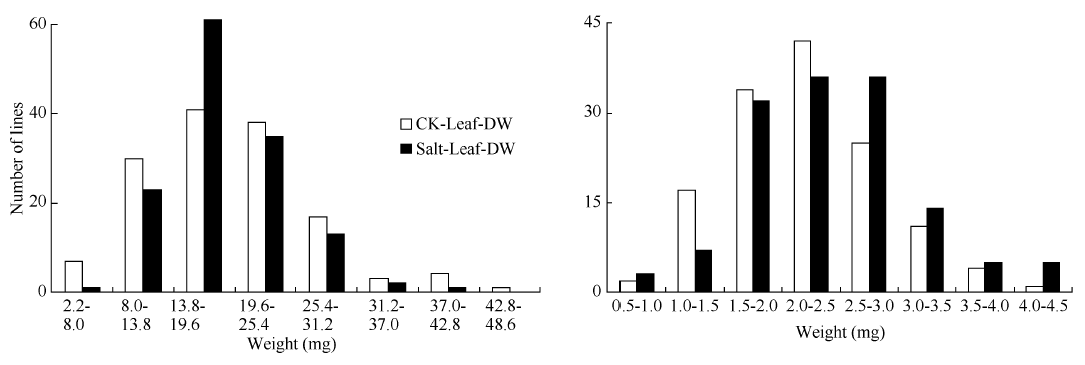
表3 油菜幼苗盐胁迫处理和对照干重、鲜重的相关系数 Table 3 Correlation coefficients between indication of dry weight and fresh weight of B. napus in salt stress treatment and control
指标 Indicator
对照叶干重 CK-Leaf-DW
对照根干重 CK-Root-DW
盐胁迫叶干重 Salt-Leaf-DW
盐胁迫根干重 Salt-Root-DW
对照叶鲜重 CK-Leaf-FW
盐胁迫叶鲜重 Salt-Leaf-FW
对照根鲜重 CK-Root-FW
对照根干重 CK-Root-DW
0.749* *
叶干重 Salt-Leaf-DW
0.205*
0.165
根干重 Salt-Root-DW
0.052
0.041
0.771* *
对照叶鲜重 CK-Leaf-FW
0.957* *
0.740* *
0.246* *
0.057
叶鲜重 Salt-Leaf-FW
0.120
0.027
0.487* *
0.429* *
0.129
对照根鲜重 CK-Root-FW
0.858* *
0.789* *
0.162
0.037
0.843* *
0.080
根鲜重 Salt-Root-FW
0.087
0.067
0.870* *
0.852* *
0.095
0.380* *
0.078
* * and * denote significant correlation at the 0.01 and 0.05 probability levels, respectively. * * 和* 分别代表在0.01和0.05显著水平。
表3 油菜幼苗盐胁迫处理和对照干重、鲜重的相关系数 Table 3 Correlation coefficients between indication of dry weight and fresh weight of B. napus in salt stress treatment and control
表4 利用复合区间作图法检测油菜幼苗干重、鲜重在正常环境下的QTL Table 4 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in normal environment
性状 Trait
数量性状位点 QTL
染色体 Chr.
标记区间 Marker interval (cM)
加性效应 Additive
贡献率 R2 (%)
阈值 LOD
置信区间 Confidence interval
对照叶干重CK-Leaf-DW
qCKDLA01
A01
SNP2668A01-SNP2079A01
-27.55
9.44
4.67
92.50-96.20
qCKDLA10
A10
SNP23037A10-SNP23245A10
-25.91
8.43
4.49
60.00-65.60
qCKDLC08
C08
SNP30078C08-SNP30068C08
31.24
12.19
6.28
35.50-36.70
对照根干重CK-Root-DW
qCKDRA01
A01
SNP2668A01-SNP1348A01
-2.13
9.13
3.34
92.50-94.60
对照叶鲜重CK-Leaf-FW
qCKWLA01
A01
SNP2668A01-SNP2079A01
-664.61
13.38
6.34
92.50-96.20
qCKWLA10
A10
SNP22233A10-SNP23011A10
-493.58
7.08
3.81
57.50-60.00
qCKWLC08-1
C08
SNP30088C08-SNP30043C08
715.82
15.08
6.58
35.30-36.80
qCKWLC08-2
C08
SNP30060-SNP30034C08
701.67
14.91
6.67
37.50-38.10
qCKWLC08-3
C08
SNP31434-SNP30196
599.27
11.66
5.42
26.70-31.80
qCKWLC08-4
C08
SNP30229C08-SNP30122C08
531.73
7.56
3.36
42.30-47.20
对照根鲜重CK-Root-FW
qCKWRA08-1
A08
SNP18210A08-SNP18201A08
32.33
8.84
3.28
40.40-41.70
qCKWRA08-2
A08
SNP12599A05-SNP18516A08
-28.81
8.62
3.59
12.50-13.50
qCKWRA08-3
A08
SNP18508A08-SNP18387A08
-29.86
8.00
4.48
14.90-26.80
表4 利用复合区间作图法检测油菜幼苗干重、鲜重在正常环境下的QTL Table 4 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in normal environment
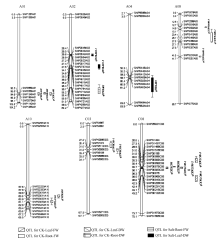
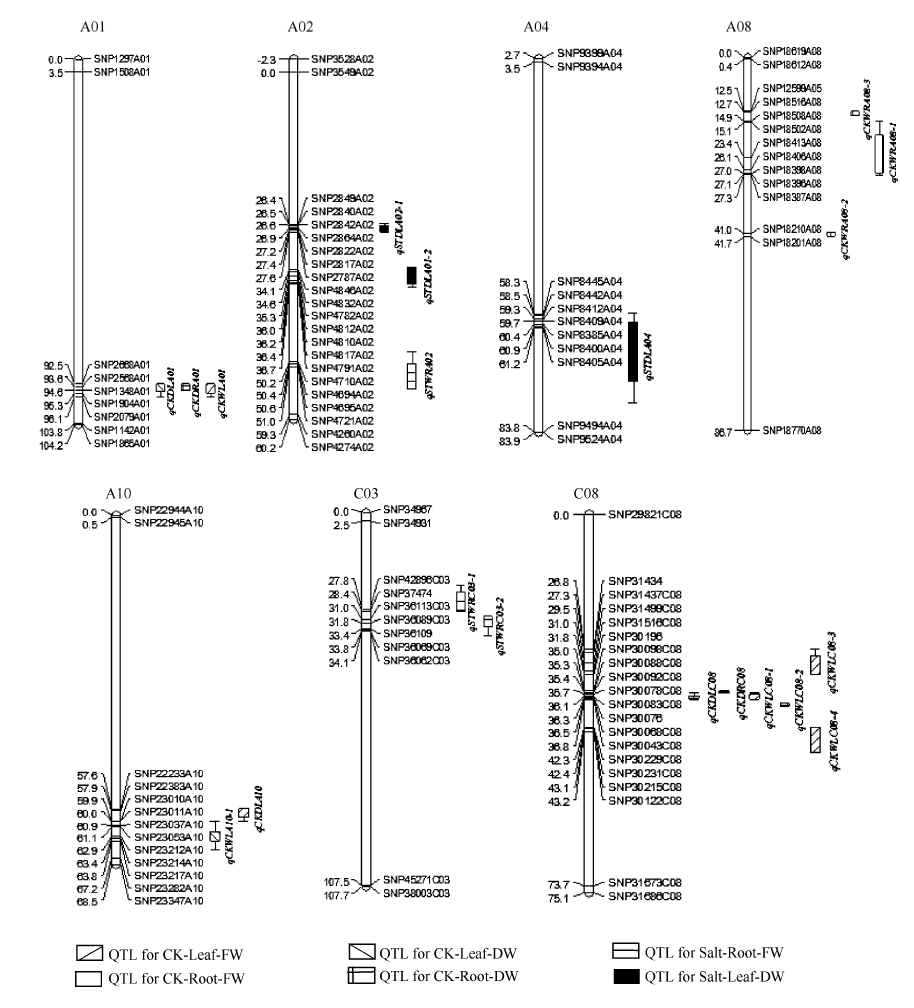
图3 甘蓝型油菜幼苗盐胁迫处理下干重、鲜重QTL在连锁群上的分布情况 QTL的染色体上仅展示了QTL区段的标记及染色体两端各2个标记。Fig. 3 Putative QTL locations of dry weight and fresh weight in salt stress treatment on the SNP genetic map Showing the markers in the QTL confidence intervals, along with the terminal two markers at each end of the QTL containing chromosomes.
表5 利用复合区间作图法检测到油菜幼苗干重、鲜重在盐胁迫环境下的QTL Table 5 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in salt stress environment
性状 Trait
数量性状位点 QTL
染色体 Chr.
标记区间 Marker interval (cM)
加性效应 Additive
贡献率 R2 (%)
阈值 LOD
置信区间 Confidence interval
盐胁迫叶干重 Salt-Leaf-DW
qSTDLA02-1
A02
SNP2849A02-SNP2787A02
-19.97
11.75
4.58
26.30-27.80
qSTDLA02-2
A02
SNP4846A02-SNP4791A02
-15.65
7.16
3.31
33.70-37.30
qSTDLA04
A04
SNP8445A04-SNP8405A04
-26.14
11.60
4.66
58.00-77.40
盐胁迫根鲜重 Salt-Root-FW
qSTWRA02
A02
SNP4046A02-SNP11915A05
-26.77
7.16
3.21
48.50-55.00
qSTWRC03-1
C03
SNP39078C03-SNP37474
39.53
16.15
4.47
20.90-28.40
qSTWRC03-2
C03
SNP36113C03-SNP36062C03
38.67
15.60
4.59
29.90-35.30
表5 利用复合区间作图法检测到油菜幼苗干重、鲜重在盐胁迫环境下的QTL Table 5 Putative QTL detected by composite interval mapping for dry weight and fresh weight of B. napus in salt stress environment
表6 甘蓝型油菜基因组中QTL置信区间候选基因与拟南芥盐胁迫相关基因的比对 Table 6 Alignment of candidate genes in QTL confidence interval in B. napus with the salt related genes in A. thaliana
数量性状座位 QTL
盐胁迫相关基因 Genes related to salt stress inA. thaliana
性状 Trait
名称 Name
物理区间 Physical interval
预测基因 Gene prediction
基因名 Gene name
登录号 Accessions number
E值 E-value
盐胁迫叶干重 Salt-Leaf-DW
qSTDLA02-1
11237912-10815211
BnaA02g18520D
SKIP/SNW
AT1G77180
E-137
qSTDLA02-2
8466214-8090196
BnaA02g14680D
ATRPK1/RPK1
AT1G69270
0
qSTDLA02-2
8466214-8090196
BnaA02g14490D
ATHVA22C
AT1G69700
2E-52
qSTDLA02-2
8466214-8090196
BnaA02g14430D
ATHB29/ZFHD1
AT1G69600
E-138
qSTDLA04
11329756-10914108
Not found
ATGRDP2
AT4G37900
5E-25
盐胁迫根鲜重 Salt-Root-FW
qSTWRC03-1
53543616-51523070
BnaC03g64020D BnaC03g64030D
ATCPL1/SHI4
AT4G21670
E-141
qSTWRC03-1
53543616-51523070
BnaC03g63430D BnaC03g63440D
CHIC
AT4G19810
E-167
qSTWRC03-1
53543616-51523070
BnaC03g62830D BnaC03g62840D
ATCESA8/LEW2
AT4G18780
4E-35
表6 甘蓝型油菜基因组中QTL置信区间候选基因与拟南芥盐胁迫相关基因的比对 Table 6 Alignment of candidate genes in QTL confidence interval in B. napus with the salt related genes in A. thaliana
表7 Table 7 表7(Table 7)
表7 筛选的在拟南芥中盐胁迫相关候选基因功能 Table 7 Functions of the screened candidate genes related to salt stress in A. thaliana
名称 Name
基因 Gene
基因功能 Gene function
参考文献 Reference
qSTDLA02-1
SKIP/SNW
编码一个假定的转录因子, 在酵母中表现转录活性, 参与到对脱落酸、盐等渗透性胁迫反应中。 Encodes a putative transcriptional factor. Shows transcriptional activator activity in yeast. Involved in response to abscisic acid, salt and osmotic stress.
RPK1是一个富含亮氨酸的受体酶, 位于离子膜中, 由脱落酸、脱水、高盐、低温等引起, 和其他植物激素无关。 RPK1 is a leucine-rich receptor-like kinase located in the plasma membrane which is upregulated by abscisic acid, dehydration, high salt, low temperature, but not by other plant hormones.
编码 ZFHD1, 它是锌指同源转录因子家族的一个成员, 其表达是由干旱、高盐和脱落酸引起的。 Encodes ZFHD1, a member of the zinc finger homeodomain transcriptional factor family. Expression of ZFHD1 is induced by drought, high salinity and abscisic acid.
编码未知功能的一种蛋白质, 参与植株生长和发育的许多方面以及应对盐胁迫。 Protein of unknown function that expressed in many plant tissues and is involved in many aspects of plant growth and development as well as response to salt stress.
编码的蛋白质在体外培养具有先天性磷酸酶活性。突变体对脱落酸, 低温和氯化钠十分敏感。 Encodes a protein that exhibits innate phosphatase activity in vitro. Mutants exhibit hyper responsiveness to ABA, cold, and NaCl.
反转录PCR实验表明, ChiC的转录水平在应对脱落酸、茉莉酸和氯化钠压力时增加。 RT-PCR experiments demonstrate that ChiC transcript levels are increased in response to abscisisc acid, jasmonic acid, and NaCl stress.
编码一个纤维素合酶家族的成员, 其参与次生细胞壁的生物合成。突变体有异常木质部形成, 会降低纤维素含量、增强干旱和渗透压力胁迫。 Encodes a member of the cellulose synthase family involved in secondary cell wall biosynthesis. Mutants have abnormal xylem formation, reduced cellulose content, and enhanced drought and osmotic stress tolerance.
杨真, 王宝山. 中国盐渍土资源现状及改良利用对策. 山东农业科学, 2015, 47(4): 125-130YangZ, Wang BS. Present status of saline soil resources and countermeasures for improvement and utilization in China. Shand ong Agric Sci, 2015, 47(4): 125-130 (in Chinese with English abstract)[本文引用:1]
[2]
QadirM, GhafoorA, MurtazaG. Amelioration strategies for saline soils: a review. Land Degrad Devel, 2000, 11: 501-521[本文引用:1]
[3]
沈金雄, 傅廷栋. 我国油菜生产、改良与食用油供给安全. 中国农业科技导报, 2011, 13(1): 1-8Shen JX, Fu TD. Rapeseed production, improvement and edible oil supply in China. J Agric Sci Technol, 2011, 13(1): 1-8 (in Chinese with English abstract)[本文引用:1]
[4]
陈宗金, 蔡士宾, 杨继书, 张巧凤, 吴纪中, 蒋彦婕, 颜伟, 吴小有. 主要农作物芽期耐盐性比较研究. 农业科学, 2012, 2(4): 59-65Chen ZJ, Cai SB, Yang JS, Zhang QF, Wu JZ, Jiang YJ, YanW, Wu XY. Comparison of salinity tolerance among main crops at germination stage. Hans J Agric Sci, 2012, 2(4): 59-65 (in Chinese with English abstract)[本文引用:1]
[5]
王佳丽, 黄贤金, 钟太洋, 陈志刚. 盐碱地可持续利用研究综述. 地理学报, 2011, 66: 673-684Wang JL, Huang XJ, Zhong TY, Chen ZG. Review on Saline-alkali land sustainable utilization research. Acta Geograph Sin, 2011, 66: 673-684 (in Chinese with English abstract)[本文引用:1]
[6]
易斌, 陈伟, 马朝芝, 傅廷栋, 涂金星. 甘蓝型油菜产量及相关性状的QTL分析. 作物学报, 2006, 32: 676-682YiB, ChenW, Ma CZ, Fu TD, Tu JX. Mapping of quantitative trait loci for yield and yield components in Brassica napus L. Acta Agron Sin, 2006, 32: 676-682 (in Chinese with English abstract)[本文引用:1]
[7]
YangP, ShuC, ChenL, Xu JS, Wu JS, Liu KD. Identification of a major QTL for silique length and seed weight in oilseed rape (Brassica napus L. ). Theor Appl Genet, 2012, 125: 285-296[本文引用:1]
[8]
孙美玉. 甘蓝型油菜含油量QTLs定位及候选基因筛选. 中国农业科学院博士论文, 湖北武汉, 2012. pp 51-54Sun MY. Mapping of QTLs and Screening of Cand idate Genes for Oil Content in Brassica napus. PhD Dissertation of Chinese Academy of Agricultural Sciences, Wuhan, China, 2012. pp 51-54 (in Chinese with English abstract)[本文引用:1]
[9]
YangM, Ding GD, ShiL, FengJ, Xu FS, Meng JL. Quantitative trait loci for root morphology in response to low phosphorus stress in Brassica napus. Theor Appl Genet, 2010, 121: 181-193[本文引用:1]
[10]
张凤启, 刘越英, 程晓辉, 童超波, 董彩华, 唐敏强, 黄军艳, 刘胜毅. 利用高密度SNP标记定位甘蓝型油菜株高QTL. 中国油料作物学报, 2014, 36: 695-700Zhang FQ, Liu YY, Cheng XH, Tong CB, Dong CH, Tang MQ, Huang JY, Liu SY. QTL mapping of plant height using high density SNP markers in Brassica napus. Chin J Oil Crop Sci, 2014, 36: 695-700 (in Chinese with English abstract)[本文引用:1]
[11]
YıldızM, AkçalıN, TerziH. Proteomic and biochemical responses of canola (Brassica napus L. ) exposed to salinity stress and exogenous lipoic acid. J Plant Physiol, 2015, 179: 90-99[本文引用:1]
[12]
刘国红, 姜超强, 刘兆普, 梁明祥, 殷祥贞, 郑青松. 盐胁迫对油菜幼苗生长和光合特征的影响. 生态与农村环境学报, 2012, 28(2): 157-164Liu GH, Jiang CQ, Liu ZP, Liang MX, Yin XZ, Zheng QS. Effects of salt stress on growth and photosynthetic traits of Canola seedlings. J Ecol Rural Environ, 2012, 28(2): 157-164 (in Chinese with English abstract)[本文引用:1]
[13]
郑青松, 刘海燕, 隆小华, 刘兆普, 牛丹丹, 高影影. 盐胁迫对油菜幼苗离子吸收和分配的影响. 中国油料作物学报, 2010, 32: 65-70Zheng QS, Liu HY, Long XH, Liu ZP, Niu DD, Gao YY. Effects of salt stress on ionic absorption and distribution of rapeseed seedlings. Chin J Oil Crop Sci, 2010, 32: 65-70 (in Chinese with English abstract)[本文引用:1]
[14]
LiZ, Mei SF, MeiZ, Liu XL, Fu TD, Zhou GS, Tu JX. Mapping of QTL associated with waterlogging tolerance and drought resistance during the seedling stage in oilseed rape (Brassica napus). Euphytica, 2014, 197: 341-353[本文引用:1]
[15]
荐红举, 肖阳, 李加纳, 马珍珍, 魏丽娟, 刘列钊. 利用SNP遗传图谱定位盐、旱胁迫下甘蓝型油菜种子发芽率的QTL. 作物学报, 2014, 40: 629-635Jian HJ, XiaoY, Li JN, Ma ZZ, Wei LJ, Liu LZ. QTL mapping for germination percentage under salinity and drought stresses in Brassica napus L. using a SNP genetic map. Acta Agron Sin, 2014, 40: 629-635 (in Chinese with English abstract)[本文引用:3]
[16]
Moursi Y SS. Genetic Mapping of QTL Controlling Salt Tolerance and Glucosinolates in Brassica napus and Brassica oleracea. PhD Dissertation of Georg-August-University, Germany, Göttingen, 2014. pp 73-78[本文引用:2]
[17]
Liu LZ, Qu CM, WittkopB, YiB, XiaoY, He YJ, Snowdon RJ, Li JN. High-Density SNP Map for Accurate Mapping of Seed Fibre QTL in Brassica napus L. PLoS One, 2013, 8: e83052. DOI: DOI:10.1371/journal.pone.0083052[本文引用:2]
[18]
WangS, Basten CJ, Zeng Z B. Windows QTL Cartographer. Ver. 2. 5 [computer program] Department of Statistics, North Carolina State University, Raleigh, NC, 2006. http://statgen.ncsu.edu/qtlcart/WQTLCart.htm[本文引用:1]
[19]
Land er ES, BotsteinD. Mapping mendelian factors underlying quantitative traits using RFLP linkage maps. Genetics, 1989, 121: 185-199[本文引用:1]
[20]
Mccouch SR, Cho YG, YanoM, PaulE, BlinstrubM, MorishimaH, KinoshitaT. Report on QTL nomenclature. Rice Genet Newslett, 1997, 14: 11-13[本文引用:1]
[21]
ChalhoubB, DenoeudF, Liu SY, Isobe IA, Tang HB, Wang XY, ChiquetJ, BelcramH, Tong CB, SamansB, CorréaM, Silva CD, JustJ, FalentinC, Koh SH, Clainche IL, BernardM, BentoP, NoelB, LabadieK, AlbertiA, CharlesM, ArnaudD, GuoH, DaviaudC, AlameryS, JabbariK, Zhao MX, Edger PP, ChelaifaH, TackD, LassalleG, MestiriI, SchnelN, Paslier ML, Fan GY, RenaultV, Bayer PE, Golicz AA, ManoliS, Lee TH, Thi V H D, Chalabi S, Hu Q, Fan C C, Tollenaere R, Lu Y H, Battail C, Shen J X, Sidebottom C H. D, Wang X F, Canaguier A, Chauveau A, Bérard A, Deniot G, Guan M, Liu Z S, Sun F M, Lim Y P, Lyons E, Christopher D, Town, Ian B, Wang X W, Meng J L, Ma J X, Pires J C, Graham J, King, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y M, Hua W, Sharpe A G, Paterson A H, Guan C Y, Wincker P. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science, 2014, 345: 950-953[本文引用:2]
[22]
Feng JL, Li JJ, Gao ZX, Lu YR, Yu JY, ZhengQ, Yan SN, Zhang WJ, HeH, Ma LG, Zhu ZG. SKIP confers osmotic tolerance during salt stress by controlling alternative gene splicing in Arabidopsis. Mol Plant, 2015, 87: 1038-1052[本文引用:1]
[23]
Hong SW, Jon JH, Kwak JM, Nam HG. Identification of a receptor-like protein kinase gene rapidly induced by abscisic acid, dehydration, high salt, and cold treatments in Arabidopsis thaliana. Plant Physiol, 1997, 113: 1203-1212[本文引用:1]
[24]
Woei-JiunG, Tuan-Hua D H. An abscisic acid-induced protein, HVA22, inhibits gibberellin-mediated programmed cell death in cereal aleurone cells. Plant Physiol, 2008, 148: 1182-1182[本文引用:1]
[25]
ElfvingN, DavoineaC, BenllochbR, BlombergaJ, BrännströmaK, MüllercD, NilssondA, UlfstedtdM, RonnedH, WingslebG, NilssonbO, BjörklundaS. The Arabidopsis thaliana Med25 mediator subunit integrates environmental cues to control plant development. Proc Natl Acad Sci USA, 2011, 108: 8245-8250[本文引用:1]
[26]
Ortega-Amaro MA, Rodriguez-Hernand ez AA, Rodriguez- KesslerM, Hernand ez-LuceroE, Rosales-MendozaS, Ibañez- SalazarA, Delgado-SanchezP, Jimenez-Bremont JF. Overexpression of AtGRDP2, a novel glycine-rich domain protein, accelerates plant growth and improves stress tolerance. Front Plant Sci, 2015, 5: 782-783[本文引用:1]
[27]
Seifert GJ, XueH, AcetT. The Arabidopsis thaliana Fasciclin like arabinogalactan protein 4 gene acts synergistically with abscisic acid signalling to control root growth. Ann Bot, 2014, 114: 1125-1133[本文引用:1]
[28]
OhnumaT, NumataT, OsawaT, MizuharaM, LampelaO, Juffer AH, SkriverK, FukamizoT. A class V chitinase from Arabidopsis thaliana: gene responses, enzymatic properties, and crystallographic analysis. Planta, 2011, 234: 123-137[本文引用:1]
MunnsR, TesterM. Mechanisms of salinity tolerance. Annu Rev Plant Biol, 2008, 59: 651-681[本文引用:1]
[31]
AshrafM, McNeilly T. Salinity tolerance in Brassica oilseeds. Crit Rev Plant Sci, 2004, 23: 157-174[本文引用:1]
[32]
TunçtürkM, TunçtürkR, YildirimB, ÇiftçiV. Changes of micronutrients, dry weight and plant development in canola (Brassica napus L. ) cultivars under salt stress. Afr J Biotechnol, 2011, 10: 3726-3730[本文引用:1]
[33]
Yeo AR. Molecular biology of salt tolerance in the context of whole-plant physiology. J Exp Bot, 1998, 49: 915-929[本文引用:1]
[34]
Flowers TJ, Yeo AR. Breeding for salinity resistance in crop plants: where next?Aust J Plant Physiol, 1995, 22: 875-884[本文引用:1]
[35]
OsakabeY, MizunoS, TanakaH, MaruyamaK, OsakabeK, TodakaD, FujitaY, KobayashiM, ShinozakiK, Yamaguchi- ShinozakiK. Overproduction of the membrane-bound receptor- like protein kinase 1, RPK1, enhances abiotic stress tolerance in Arabidopsis. J Biol Chem, 2010, 285: 9190-9201[本文引用:1]
[36]
Shi CC, Feng CC, Yang MM, Li JL, Li XX, Zhao BC, Huang ZJ, Ge RC. Overexpression of the receptor-like protein kinase genes AtRPK1 and OsRPK1 reduces the salt tolerance of Arabidopsis thaliana. Plant Sci, 2014, 217: 63-70[本文引用:1]
[37]
Long WH, Zou XL, Zhang XK. Transcriptome analysis of canola (Brassica napus) under salt stress at the germination stage. PLoS One, 2015, 10(2): e0116217[本文引用:1]
 , 王腾岳
, 王腾岳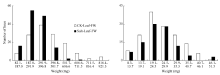


{kind=link}
{kind=link}
{kind=link}
{kind=link}