关键词:茶树花; 基因表达谱; 不育基因; 差异表达基因 Exploring Sterility Gene from Tea Plant Flower Based on Digital Gene Expression Profiling CHEN Lin-Bo1,**, XIA Li-Fei1,**, TIAN Yi-Ping1, LI Mei1, SONG Wei-Xi1, LIANG Ming-Zhi1,*, JIANG Chang-Jun2,* 1 Tea Research Institute, Yunnan Academy of Agricultural Sciences / Yunnan Engineering Research Center of Tea Germplasm Innovation and Matching Cultivation, Menghai 666201, China
2State Key Laboratory of Tea Plant Biology and Utilization, Anhui Agricultural University, Hefei 230036, China
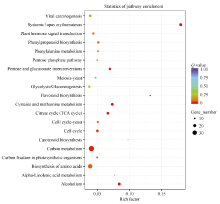
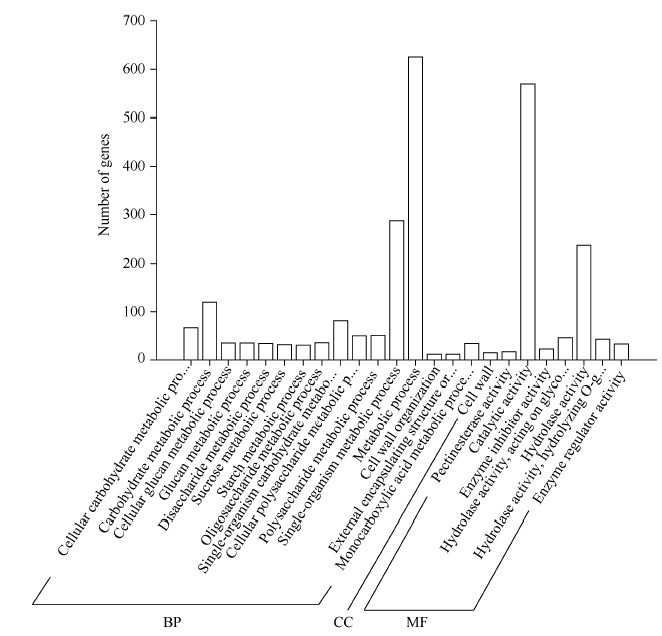
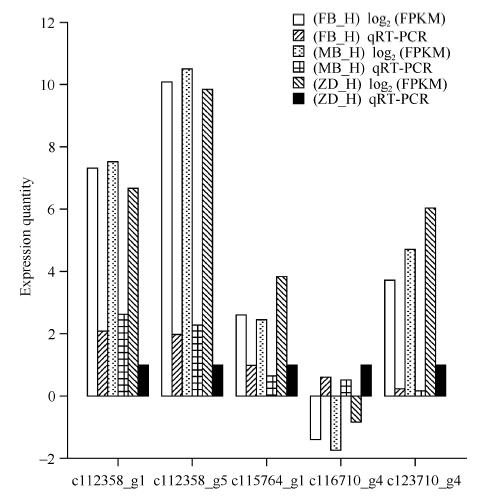
Fund:This study was supported by the National Natural Science Foundation of China (31560220, 31460216), the Personnel Training Program of Yunnan Provence (2015HB105), and the Open Fund of State Key Laboratory of Tea Plant Biology and Utilization (SKLTOF20150105) AbstractTea tree ( Camellia sinensis) blossoms every year and a lot of nutrients are consumed by itself during blooming and fruiting, resulting in decreased yield and quality of fresh tea leaf. In the present study, the flowers of Fuding Dabaicha (male parent), Foxiang 2 (female parent) and their hybrid (sterile flowers) were used to construct cDNA libraries and the gene expression profiles were analyzed by Solexa sequencing technology. A total of 1219 candidate sterility genes were identified according to their differential expression between the male/female parent and the hybrid but identical between the two parents. These sterility gene were subject to GO analysis, and metabolic process, hydrolase activity and catalytic activity were enriched. KEGG pathway analysis showed that the sterility genes were involved in biosynthesis of amino acids and carbohydrate, second any metabolism, plant hormone signal transduction, and energy metabolism. In the function group of plant hormone signal transduction, 16 annotated genes were involved in the auxin signal transduction pathway, including five up-regulated AFR genes in the hybrid and 11 down-regulated genes. Thus, we speculate that auxin signal transduction might regulate the flower sterility in tea tree. Five sterility genes were selected randomly for qRT-PCR assay and the result was consistent with that of Solexa sequencing. The candidate sterility genes found in this study are worthy of sterile mechanism research in tea plant and sterility gene screening.
Keyword:Tea plant flowers; Gene expression profiles; Sterility gene; Differentially expressed genes Show Figures Show Figures
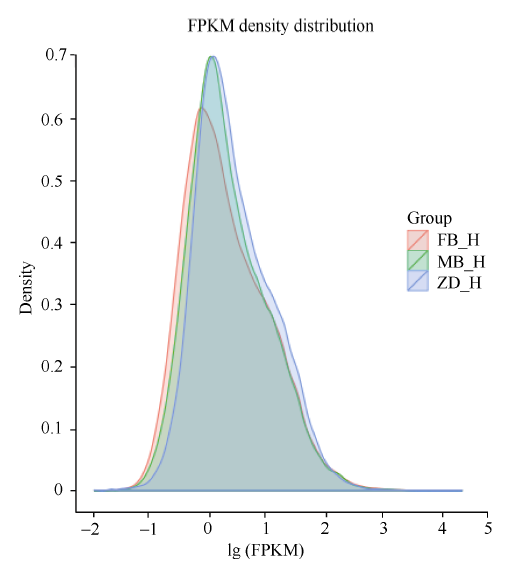
图2 不同样的基因表达水平比对 FB_H为父本花; MB_H母本花; ZD_H为不育花。Fig. 2 Comparison of genes expression of different samples FB_H is male parent flowers; MB_H is female parent flowers; ZD_H is sterility flowers.
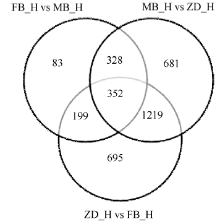
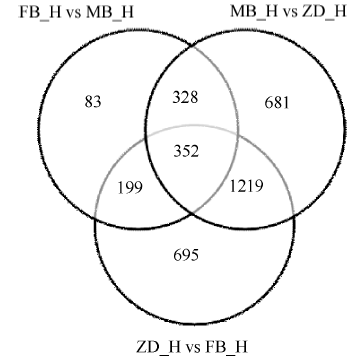
图3 差异基因维恩图 FB_H为父本花; MB_H母本花; ZD_H为不育花。Fig. 3 Venn image of differential genes FB_H is male parent flowers; MB_H is female parent flowers; ZD_H is sterility flowers.
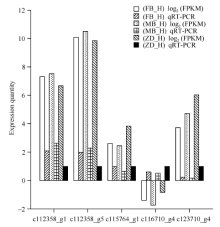
图6 Illumina HiSeq测序和qRT-PCR的表达量比对 FB_H为父本花; MB_H母本花; ZD_H为不育花。Fig. 6 Validation of Illumina HiSeq sequencing data by qRT-PCR FB_H is male parent flowers; MB_H is female parent flowers; ZD_H is sterility flowers.
黄亚辉. 浅析控制茶树生殖生长的途径. 中国茶叶, 1997, (4): 6-7Huang YH. Analyses the way of control reproductive growth in tea plant. China Tea, 1997, (4): 6-7 (in Chinese)[本文引用:1]
[2]
夏春华, 束际林. 茶树化学除花与茶叶增产. 植物杂志, 1979, (2): 6-7Xia CH, Shu JL. Chemical flowers removing and increase tea production in tea plant. Plant J, 1979, (2): 6-7 (in Chinese with English abstract)[本文引用:1]
[3]
吴淑平. 茶树营养生长与生殖生长的关系及调控方法. 中国园艺文摘, 2011, 27(5): 182-183Wu SP. The relationship and control method of vegetative growth and reproductive growth in tea plant. Chin Hort Abstr, 2011, 27(5): 182-183 (in Chinese with English abstract)[本文引用:1]
[4]
Tang RS, Zheng JC, Jin ZQ, Zhang DD, Huang YH, Chen LG, Tang RS. Possible correlation between high temperature-induced floret sterility and endogenous levels of IAA, GAs and ABA in rice (Oryza sativa L. ). Plant Growth Regul, 2008, 54: 37-43[本文引用:2]
[5]
Wan GX, Li SQ, LiW, JinW, KunW, Zhu YG. Damage of oxidative stress on mitochondria during microspores development in Honglian CMS line of rice. Plant Cell Rep, 2007, 26: 373-382[本文引用:2]
[6]
冯小磊, 范光宇, 苏旭, 宋国亮, 史高雷, 王峰, 邱风仓, 王晓明, 赵治海. 植物雄性不育生理生化研究进展. 作物杂志, 2012, (3): 6-11Feng XL, Fan GY, SuX, Song GL, Shi GL, WangF, Qiu FC, Wang XM, Zhao ZH. Advances in physiological and biochemical study on plant male sterility. Crops, 2012, (3): 6-11 (in Chinese with English abstract)[本文引用:1]
[7]
王继玥, 柯剑鸿. 植物细胞质雄性不育分子机制研究进展. 玉米科学, 2015, 23(3): 7-13Wang JY, Ke JH. Research progress on molecular mechanism of cytoplasmic male sterility in plant. J Maize Sci, 2015, 23(3): 7-13 (in Chinese with English abstract)[本文引用:1]
[8]
汤继华, 赫忠友, 陈伟程, 谭树义, 谢惠玲, 李永亮. 玉米温敏核雄性不育育性转换与内源激素的关系. 作物学报, 2003, 29: 336-338Tang JH, He ZY, Chen WC, Tan SY, Xie HL, Li YL. Relationship between fertility conversion and endogenous hormones in a maize thermo-sensitive genic male-sterile line. Acta Agron Sin, 2003, 29: 336-338 (in Chinese with English abstract)[本文引用:2]
[9]
解海岩, 蒋培东, 王晓玲, 张昭伟, 朱伟, 王学德. 棉花细胞质雄性不育花药败育过程中内源激素的变化. 作物学报, 2006, 32: 1094-1096Xie HY, Jiang PD, Wang XL, Zhang ZW, ZhuW, Wang XD. Changes of phytohormone contents in anther abortion of cytoplasmic male sterile cotton. Acta Agron Sin, 2006, 32: 1094-1096 (in Chinese with English abstract)[本文引用:1]
[10]
Yin CX, Gan LJ, NgD, Xia LK. Decreased panicle-derived indole-3-acetic acid reduces gibberellin A1 level in the uppermost internode, causing panicle enclosure in male sterile rice Zhenshan 97A. J Exp Bot, 2012, 58: 2441-2449[本文引用:1]
[11]
任怡怡, 戴绍军, 刘炜. 生长素的运输及其在信号转导及植物发育中的作用. 生物技术通报, 2012, (3): 9-16Ren YY, Dai SJ, LiuW. Auxin transport and its roles in signal transduction and plant development. Biotechnol Bull, 2012, (3): 9-16 (in Chinese with English abstract)[本文引用:2]
[12]
Shi CY, YangH, Wei CL, YuO, Zhang ZZ, Jiang CJ, SunJ, Li YY, ChenQ, XiaT, Wan XC. Deep sequencing of the Camellia sinensis transcriptome revealed cand idate genes for major metabolic pathways of tea-specific compounds. BMC Genom, 2011, 12: 131[本文引用:1]
[13]
Alagna FD, AgostinoN, TorchiaL, ServiliM, RaoR, PietrellaM, GiulianoG, Chiusano ML, BaldoniL, PerrottaG. Comparative 454 pyrosequencing of transcripts from two olive genotypes during fruit development. BMC Genom, 2009, 10: 399[本文引用:1]
[14]
史成颖, 宛晓春, 江昌俊, 孙俊. 提取高质量茶树总RNA的方法研究. 安徽农业大学学报, 2007, 34: 360-363Shi CY, Wan XC, Jiang CJ, SunJ. Method for high quality total RNA isolation from tea plant [Camellia sinensis (L. ) O. Kuntze]. J Anhui Agric Univ, 2007, 34: 360-363 (in Chinese with English abstract)[本文引用:2]
[15]
江昌俊, 王朝霞, 李叶云. 茶树中提取总RNA的研究. 茶叶科学, 2000, 20(1): 27-29Jiang CJ, Wang ZX, Li YY. Studies on isolation of general RNA from tea plant. J Tea Sci, 2000, 20(1): 27-29 (in Chinese with English abstract)[本文引用:2]
[16]
MortazaviA, Williams BA, McCue K, Schaeffer L, Wold B Mapping and quantifying mammalian transcriptomes by RNA-seq. Natl Methods, 2008, 5: 621-628[本文引用:1]
[17]
WangG, Zhu QG, Meng QW, Wu CG. Transcript profiling during salt stress of young cotton (Gossypium hirsutum) seedlings via Solexa sequencing. Acta Physiol Plant, 2012, 34: 107-115[本文引用:1]
[18]
Yan ZX, Shao JZ, DingY. Catalytic assays in blue native gels revealed normal ATPase but deficient NADH dehydrogenase activity in ZidaoA CMS line of rice. Acta Physiol Plant, 2011, 33: 2477-2484[本文引用:1]
[19]
TheissenG. Development of floral organ identity: stories from the MADS house. Curr Opin Plant Biol, 2001, 4: 75-85[本文引用:1]
[20]
HeijmansK, MorelP, Vand enbusscheM. MADS-box genes and floral development: the dark side. J Exp Bot, 2012, 63: 5397-5404[本文引用:2]
[21]
BenkováE, MichniewiczM, SauerM, FrimlJ. Local, efflux-dependent auxin gradients as a common module for plant organ formation. Cell, 2003, 115: 591-602[本文引用:1]
[22]
Bohn-CourseauI. Auxin: a major regulator of organogenesis. Plant Biol Pathol, 2010, 333: 290-296[本文引用:1]
[23]
刘振华, 于延冲, 向凤宁. 生长素响应因子与植物的生长发育. 遗传, 2011, 33: 1335-1346Liu ZH, Yu YC, Xiang FN. Auxin response factors and plant growth and development, Hereditas (Beijing), 2011, 33: 1335-1346 (in Chinese with English abstract)[本文引用:1]
GuilfoyleT. Plant biology: sticking with auxin. Nature, 2007, 446: 621-622[本文引用:1]
[26]
TanX, Calderon-Villalobos L I, Sharon M, Zheng C X, Robinson C V, Estelle M, Zheng N. Mechanism of auxin perception by the TIR1 ubiquitin ligase. Nature, 2007, 446: 640-645[本文引用:1]
 , 夏丽飞
, 夏丽飞



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}