关键词:水稻; 早衰; 基因定位; PSLS1基因; 谷氨酸合酶(Fd-GOGAT) Gene Mapping and Cloning of a Premature Leaf Senescence Mutant psls1 in Rice HUANG Ya-Min1,**, ZHU Shan-Shan1,**, ZHAO Zhi-Chao1,**, PU Zhi-Gang2, LIU Tian-Zhen1, LUO Sheng1, ZHANG Xin1,* 1 National Key Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2 Biotechnology Institute of Nuclear Technology, Sichuan Academy of Agricultural Sciences, Chengdu 610066 , China
Fund:This study was supported by the National Natural Science Foundation of China (91535302) and the Major Project of China on New Varieties of GMO Cultivation (2015ZX08010-004) AbstractLeaf early senescence directly reduces its photosynthetic capacity, decreasing crop yield and grain quality. It is of great importance to identify novel mutants and characterize their molecular and physiological mechanisms. In this paper, we reported gene mapping and cloning of a psls1 mutant (premature senescence leaf with spots) in rice. The mutant showed early senescence symptoms, including decreased chlorophyll content, over-accumulated H2O2, yellow-withered leaf and increased dead cell from bottom to top of plant after the 7-leaf stage. Moreover, the plant height, tiller number, panicle length, and the number of grains per panicle were significantly lower in psls1. We observed the degradation of chloroplast, unclean thylakoid and numerous osmiophilic granules in psls1leaves by transmission electron microscope (TEM). Genetic analysis demonstrated that the phenotype of psls1 was determined by a single recessive gene. Using genetic population derived from psls1× IRAT129, the psls1 locus was mapped in a 89 kb region flanked by markers ZS-3 and ZS-8 on chromosome 7, containing 12 putative open reading frames (ORFs). Sequence analysis revealed a single-base substitution occurred in the genomic sequence of LOC_Os07g46460, which led to a 59 bp deletion in its cDNA, and therefore, predicted LOC_Os07g46460( PSLS1), encoding a ferredoxin-dependent glutamate synthase as the candidate gene. The mRNA expression level of mutated PSLS1 decreased sharply, resulting in reduction of glutamate synthase activity and abnormal amino acid metabolism in psls1. Under low nitrogen treatment, the senescence phenotype of psls1could occur as early as at the 3-leaf stage. These results indicated that psls1 senescence phenotype might be associated with the loss of glutamate synthase activity and the abnormal amino acid metabolism.
Keyword:Rice; Leaf senescence; Gene-mapping; PSLS1 gene; Glutamate synthase (Fd-GOGAT) Show Figures Show Figures
图1 野生型和突变体psls1表型 A: 野生型(WT)和突变体psls1幼苗期表型; B: 野生型(WT)和突变体psls1分蘖初期表型(下部叶片见图C左, 上部叶片见图C右); D: 野生型(WT)和突变体psls1拔节后期表型(上部叶片见图E)。Fig. 1 Phenotype of wild type and psls1 mutant A: phenotype of WT andpsls1 at seedling stage; B: phenotype of WT andpsls1 at early tillering stage (lower leaves and upper leaves are displayed in C); D: phenotype of WT andpsls1at late elongation stage (upper leaves are displayed in E).
表1 Table 1 表1(Table 1)
表1 野生型与psls1突变体农艺性状比较 Table 1 Comparison on agronomic traits between wild type and psls1 mutant
农艺性状 Agronomic trait
野生型 Wild type
突变体 psls1
株高 Plant height (cm)
108.15± 2.54
86.76± 1.76* *
分蘖数 Tiller number
11.00± 1.15
8.30± 0.48* *
结实率 Seed-setting rate
84.66± 0.06
78.80± 0.04
穗长 Main panicle length (cm)
29.30± 0.25
24.90± 0.48* *
每穗实粒数 Grain number per panicle
289.60± 15.04
164.20± 10.85* *
初级枝梗数 No. of primary branch
11.60± 0.55
10.60± 0.55*
二级枝梗数 No. of secondary branch
54.80± 3.19
28.00± 2.91* *
千粒重 1000-grain weight
28.80± 0.46
27.10± 0.64* *
* * represents significance at P < 0.01, * represents significance at P < 0.05. * * 表示在0.01水平上差异显著, * 表示在0.05水平上差异显著。
表1 野生型与psls1突变体农艺性状比较 Table 1 Comparison on agronomic traits between wild type and psls1 mutant
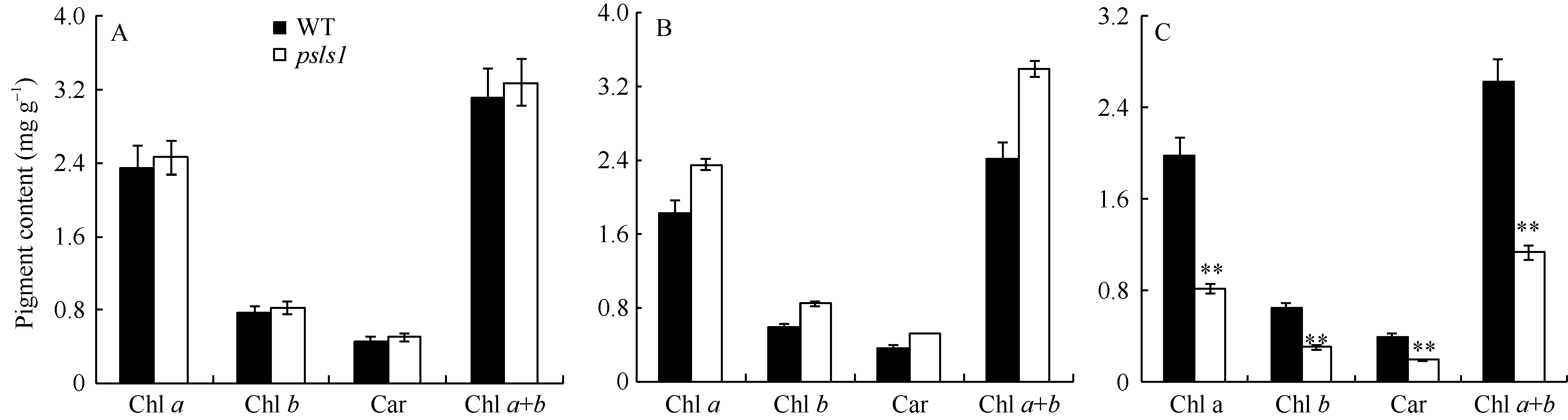
图2 野生型(WT)和突变体psls1光合色素含量 A: 野生型与突变体psls1幼苗(未出现早衰)第4片真叶光合色素含量; B: 野生型WT与突变体psls1 (严重早衰表型)绿色心叶光合色素含量; C: 野生型与突变体psls1抽穗前(严重早衰表型)倒三叶光合色素含量。Fig. 2 Photosynthetic pigment content of wild type (WT) and psls1 mutant A: pigment content of the fourth leaf in seedling (non chlorisis); B: pigment content of the green leaf before heading stage (serious chlorisis); C: pigment content of the 3rd leaf from the top before heading stage (serious chlorisis).
图3 野生型(WT)与突变体psls1组织化学染色及过氧化氢检测 A: 分蘖后期野生型(绿叶)和突变体psls1 (黄化叶)DAB染色; B: 野生型(绿叶)和突变体psls1 (黄化叶)台酚蓝染色; C: 抽穗前期心叶及倒三叶过氧化氢含量。Fig. 3 Histochemical staining and detection on hydrogen peroxide content of wild type (WT) and psls1 mutant A: DAB staining of wild type (green leaf) and psls1(chlorotic leaf); B: Trypan Blue staining of wild type (green leaf) and psls1 (chlorotic leaf); C: H2O2 content of green leaf and chlorotic leaf (the 3rd leaf from the top) before heading stage.
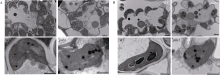
图5 野生型WT与突变体psls1叶肉细胞超微结构 A: 野生型WT与突变体psls1上部叶片叶绿体结构(psls1微黄); B: 野生型WT与突变体psls1下部叶片叶绿体结构(psls1严重黄化)。Fig. 5 Ultrastructure of mesophyll cell in mutant psls1 and the wild type A: chloroplast structure of upper leaves in wild type and psls1 (psls1slightly yellowing); B: chloroplast structure of lower leaves in wild type and psls1 (psls1serious chlorisis).
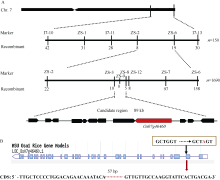
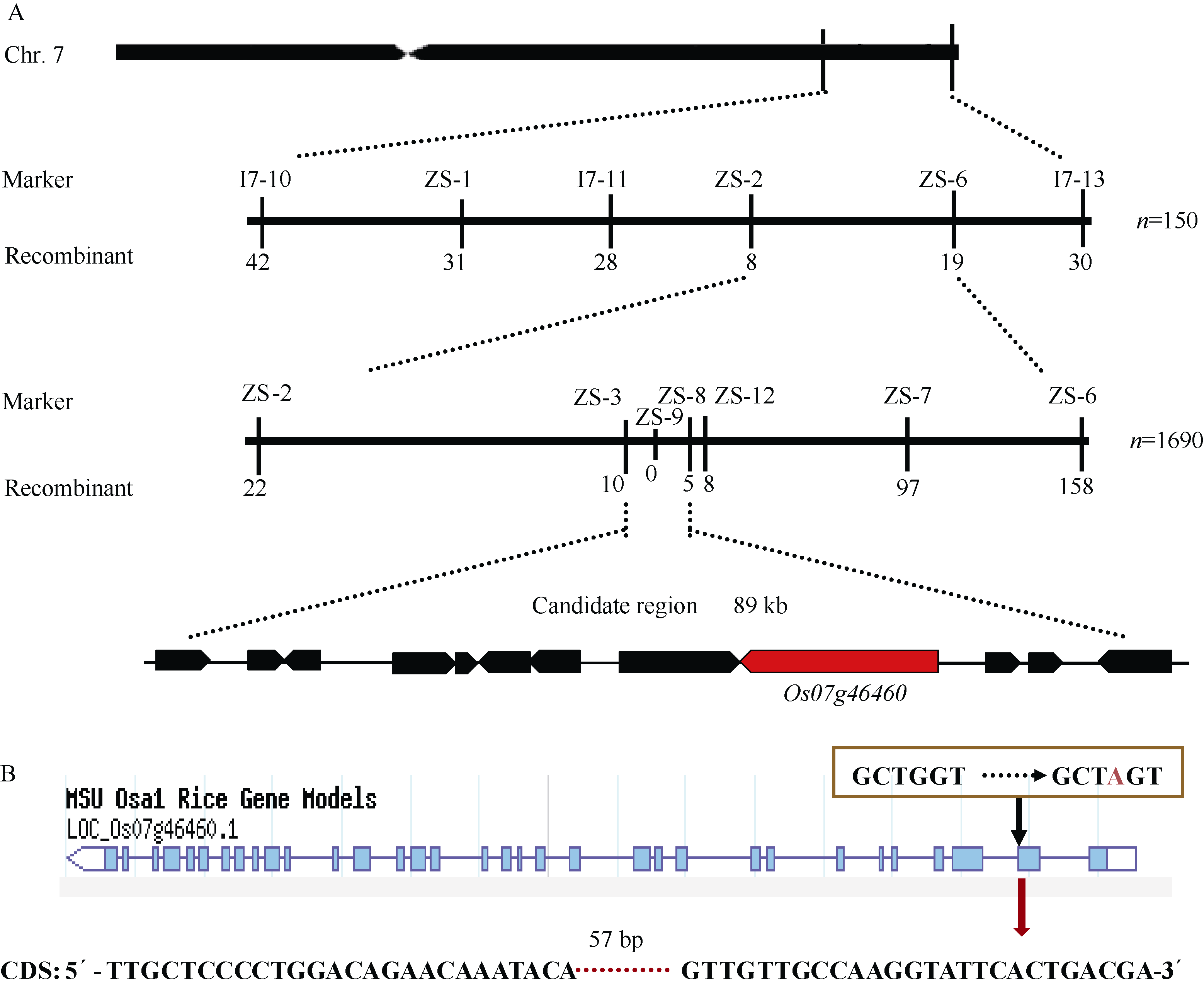
图6PSLS1的图位克隆 A: PSLS1基因的精细定位(横线上方的标记表示精细定位中用的SSR和Indel标记, 下方的数字表示该标记下的重组交换株数)。B: 定位区间基因LOC_Os07g46460基因组G→ A的改变导致编码区存在57 bp碱基缺失。Fig. 6 Map-based cloning of PSLS1gene A: fine mapping of PSLS1(the numerals above bar represent SSR and Indel markers used and the numerals below bar represent recombinants); B: the sequence variations in candidate region, the mutation of single base in LOC_Os07g46460leads to base delection of 57 bp in CDS.
表2 Table 2 表2(Table 2)
表2 候选区间内基因预测 Table 2 Putative genes in the delimited genomic region
基因 Gene locus
注释功能 Putative function
LOC_Os07g46380
Glycosyltransferase, putative, expressed
LOC_Os07g46390
TRAF-type zinc finger domain-containing protein 1, putative, expressed
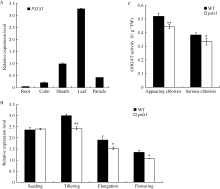
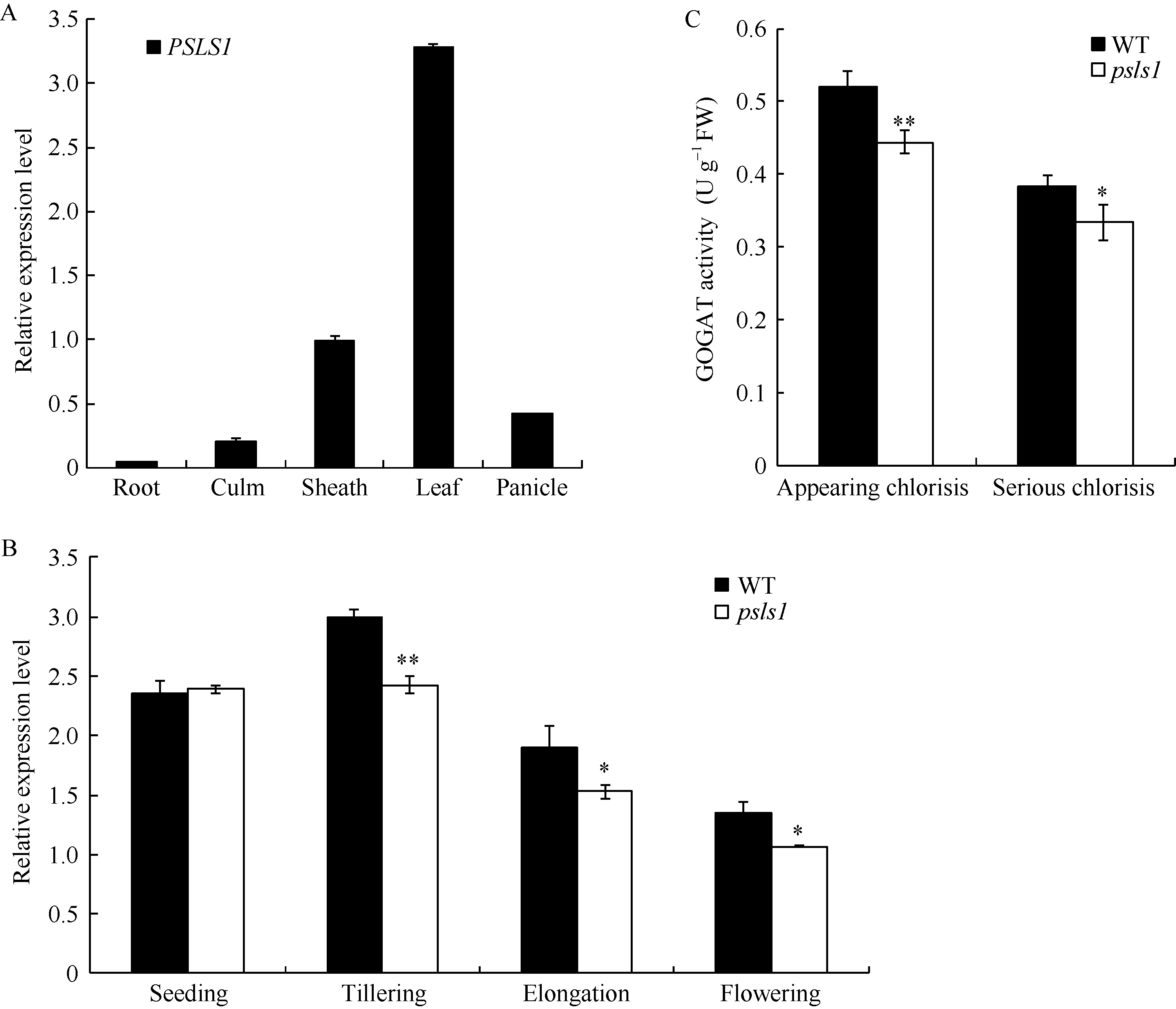
图7PSLS1表达量分析和谷氨酸合酶活性测定 A: PSLS1组织特异性表达分析; B: 不同生长时期PSLS1表达量; C: psls1刚出现早衰表型(appearing chlorisis)和早衰表型严重期(serious chlorisis)谷氨酸合酶活性。Fig. 7 Relative expression level of PSLS1and GOGAT activity A: tissue-specific expression of PSLS1; B: expression of PSLS1 in different periods; C: GOGAT activity of wild type and psls1(in appearing chlorisis and serious chlorisis stages respectively).
表3 Table 3 表3(Table 3)
表3 野生型与psls1叶片中游离氨基酸含量 Table 3 Free amino acid content in leaves of wild-type and psls1mutant (μ g g-1)
氨基酸 Amino acid
野生型 WT
突变体 psls1
氨基酸 Amino acid
野生型 WT
突变体 psls1
PSer
50.7± 0.45
91.4± 0.15* *
Leu
21.8± 0.20
18.3± 0.11
Thr
71.7± 0.77
94.0± 0.75* *
Tyr
18.8± 0.95
26.8± 0.64*
Ser
185.4± 2.30
42.1± 0.86* *
Phe
30.0± 0.69
57.9± 0.76* *
Asn
11.9± 0.61
21.2± 0.61* *
His
12.2± 0.31
54.5± 0.39* *
Glu
1504.2± 15.37
458.6± 14.91* *
Trp
124.0± 5.80
138.5± 0.93*
Gly
34.1± 0.83
17.5± 0.19* *
Orn
8.9± 0.47
30.0± 0.19* *
Ala
479.4± 0.74
167.6± 1.24* *
Lys
17.1± 0.45
38.5± 0.57* *
Val
63.4± 0.31
27.3± 0.21* *
Arg
8.2± 0.96
23.6± 0.02* *
Ile
11.6± 0.23
0* *
Pro
53.3± 1.92
54.0± 1.50
Cys
0
68.7± 0.38* *
Data are presented as means ± SD. Three independent plants were measured to calculate the means value. * Significant at P< 0.05; * * Significant at P< 0.01. * 表示在P< 0.05水平上差异显著, * * 表示在P< 0.01水平上差异显著。
表3 野生型与psls1叶片中游离氨基酸含量 Table 3 Free amino acid content in leaves of wild-type and psls1mutant (μ g g-1)
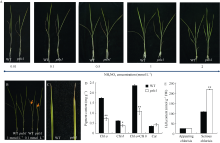
图8 低氮处理下野生型和psls1表型及生理变化 A: 不同浓度梯度氮素处理下野生型WT和突变体psls1表型(从左到右NH4NO3浓度依次为0.01 mmol L-1、0.1 mmol L-1、0.5 mmol L-1、1 mmol L-1、2 mmol L-1); B: 1 mmol L-1 (左)和0.1 mmol L-1(右) NH4NO3条件下野生型WT和突变体psls1表型(红色箭头处见图C); D: 低氮处理(0.1 mmol L-1 NH4NO3)下四叶期(第3叶)色素含量; E: 低氮处理(0.1 mmol L-1 NH4NO3)三叶期(appearing chlorisis)和四叶期(serious chlorisis)过氧化氢含量。Fig. 8 Phenotype and physiological changes of wild type and psls1 mutant under low nitrogen condition A: phenotype of wild type and psls1 mutant under different concentrations of nitrogen (NH4NO3) (concentration from left to right in turn are 0.01 mmol L-1, 0.1 mmol L-1, 0.5 mmol L-1, 1 mmol L-1, and 2 mmol L-1); B: phenotype of wild type and psls1 mutant under 1 mmol L-1 and 0.1 mmol L-1 NH4NO3 (the red arrow mark the leaf magnified in C); D: pigment content of wild type and psls1at four-leaf stage (the third leaf) under low nitrogen treatment (0.1 mmol L-1); E: H2O2 content of wild type and psls1 at three-leaf stage (appearing chlorisis) and four-leaf stage (serious chlorisis) under low nitrogen solution condition (0.1 mmol L-1).
GrbicV, Bleecker AB. Ethylene regulates the timing of leaf senescence in Arabidopsis. Plant J, 1995, 8: 595-602[本文引用:1]
[3]
Alonso JM, HirayamaT, RomanG, NourizadehS, Ecker JR. EIN2, a bifunctional transducer of ethylene and stress responses in Arabidopsis. Science, 1999, 284: 2148-2152[本文引用:1]
[4]
Li ZH, Peng JY, WenX, Guo HW. ETHYLENE-INSENSITIVE3 is a senescence-associated gene that accelerates age-dependent leaf senescence by directly repressing miR164 transcription in Arabidopsis. Plant Cell, 2013, 25: 3311-3328[本文引用:1]
[5]
Liang CZ, Wang YQ, Zhu YN, Tang JY, HuB, Liu LC, Ou SJ, Wu HK, Sun XH, Chu JF, Chu CC. OsNAP connects abscisic acid and leaf senescence by fine-tuning abscisic acid biosynthesis and directly targeting senescence-associated genes in rice. Proc Acad Natl Sci USA, 2014, 111: 10013-10018[本文引用:2]
[6]
DanismanS, van der WalF, DhondtS, WaitesR, de FolterS, BimboA, van DijkA, Muino JM, CutriL, Dornelas MC, Angenent GC, Immink R GH. Arabidopsis class I and class II TCP transcription factors regulate jasmonic acid metabolism and leaf development antagonistically. Plant Physiol, 2012, 159: 1511-1523[本文引用:1]
[7]
Rivero RM, KojimaM, GepsteinA, SakakibaraH, MittlerR, GepsteinS, BlumwaldE. Delayed leaf senescence induces extreme drought tolerance in a flowering plant. Proc Natl Acad Sci USA, 2007, 104: 19631-19636[本文引用:1]
[8]
MorrisK, MacKerness S A H, Page T, John C F, Murphy A M, Carr J P, Buchanan-Wollaston V. Salicylic acid has a role in regulating gene expression during leaf senescence. Plant J, 2000, 23: 677-685[本文引用:1]
[9]
FukaoT, YeungE, Bailey-SerresJ. The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through bormonal regulation in rice. Plant Physiol, 2012, 160: 1795-1807[本文引用:1]
[10]
Han MH, Kim CY, LeeJ, Lee SK, Jeon JS. OsWRKY42 represses OsMT1d and induces reactive oxygen species and leaf senescence in rice. Mol Cell, 2014, 37: 532-539[本文引用:1]
[11]
Rao YC, Yang YL, XuJ, Li XJ, Leng YJ, Dai LP, Huang LC, Shao GS, Ren DY, HuJ, Guo LB, Pan JW, Zeng DL. EARLY SENESCENCE1 encodes a SCAR-LIKE PROTEIN2 that affects water loss in rice. Plant Physiol, 2015, 169: 1225-1239[本文引用:1]
[12]
Gao QS, Yang ZF, ZhouY, Yin ZT, QiuJ, Liang GH, Xu CW. Characterization of an Abc1 kinase family gene OsABC1-2 conferring enhanced tolerance to dark-induced stress in rice. Gene, 2012, 498: 155-163[本文引用:1]
[13]
KusabaM, ItoH, MoritaR, IidaS, SatoY, FujimotoM, Kawasa-ki S, Tanaka R, Hirochika H, Nishimura M, Tanaka A. Rice NON-YELLOW COLORING1 is involved in light-harvesting complex II and grana degradation during leaf senescence. Plant Cell, 2007, 19: 1362-1375[本文引用:1]
[14]
Park SY, Yu JW, Park JS, Li JJ, Yoo SC, Lee NY, Lee SK, Jeong SW, SeoH, Koh HJ, Jeon JS, ParkY, Paek NC. The senescence-induced stay green protein regulates chlorophyll degradation. Plant Cell, 2007, 19: 1649-1664[本文引用:1]
[15]
YoshidaS, ItoM, NishidaI, AkiraW. Identification of a novel gene HYS /CPR5 that has a repressive role in the induction of leaf senescence and pathogen defence responses in Arabidopsis thaliana. Plant J, 2002, 29: 427-437[本文引用:1]
[16]
AkiT, KonishiM, KikuchiT, FujimoriT, YoneyamaT, YanagisawaS. Distinct modulations of the hexokinase1-mediated glucose response and hexokinase1-independent processes by HYS1/CPR5 in Arabidopsis. J Exp Bot, 2007, 58: 3239-3248[本文引用:1]
[17]
Lea PJ, Miflin BJ. Alternative route for nitrogen assimilation in higher plants. Nature, 1974, 251: 614-616[本文引用:1]
[18]
Lam HM, Coschigano KT, Oliveira IC, OliveiraM, Coruzzi G M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Annu Rev Plant Physiol Plant Mol Biol, 1996, 47: 569-593[本文引用:4]
[19]
Somerville CR, Ogren WL. Inhibition of photosynthesis in Arabidopsis mutants lacking leaf glutamate synthase activity. Nature, 1980, 286: 257-259[本文引用:1]
[20]
SuzukiA, RothsteinS. Structure and regulation of ferredoxin-dependent glutamate synthase from Arabidopsis thaliana: cloning of cDNA, expression in different tissues of wild-type and gltS mutant strains, and light induction. Eur J Biochem, 1997, 243: 708-718[本文引用:2]
[21]
Coschigano KT, Melo-OliveiraR, LimJ, Coruzzi GM. Arabidopsis gls mutants and distinct Fd-GOGAT genes: implications for photorespiration and primary nitrogen assimilation. Plant Cell, 1998, 10: 741-752[本文引用:3]
[22]
SuzukiA, VidalJ, GadalP. Glutamate synthase isoforms in rice: immunological studies of enzymes in green leaf, etiolated leaf, and root tissues. Plant Physiol, 1982, 70: 827-832[本文引用:2]
[23]
Zhao XQ, Shi WM. Expresssion analysis of the glutamine synthetase and glutamate synthase gene families in young rice (Oryza sativa) seedlings. Plant Sci, 2006, 170: 748-754[本文引用:2]
[24]
卢永恩, 罗风, 杨猛, 李香花, 练兴明. 抑制表达谷氨酸合酶基因对水稻碳氮代谢的影响. 生命科学, 2011, 41: 481-493Lu YN, LuoF, YangM, Li XH, Lian XM. Suppression of glutamate synthase genes significantly affects carbon and nitrogen metabolism in rice (Oryza sativa L. ). Sci China Life Sci, 2011, 41: 481-493 (in Chinese)[本文引用:4]
[25]
MattanaM, BiazziE, BertaniA, CoraggioI. Characterization of the Ferredoxin-Gogat gene (OsGog2 clone) expression in rice. Biol Plant, 2006, 50: 187-192[本文引用:1]
[26]
WuZ, ZhangX, HeB, DiaoL, ShengS, WangJ, GuoX, SuN, WangL, JiangL, WangC, ZhaiH, WanJ. A chlorophyll-deficient rice mutant with impaired chlorophyllide esterification in chlorophyll biosynthesis. Plant Physiol, 2007, 45: 29-40[本文引用:1]
[27]
Thordal-ChristensenH, ZhangZ, WeiY, Collinge DB. Sub-cellular localization of H2O2 in plants. H2O2 accumulation in papillaeand hypersensitive response during the barley-powdery mildew interaction. Plant J, 1997, 11: 1187-1194[本文引用:1]
DongH, Fei GL, Wu CY, Wu FQ, Sun YY, Chen MJ, Ren YL, Zhou KN, Cheng ZJ, Wang JL, JiangL, ZhangX, Guo XP, Lei CL, SuN, Wang HY, Wan JM. A rice Virescent-yellow leaf mutant reveals new insights into the role and assenbly of plastid caseinolytic protease in higher plants. Plant Physiol, 2013, 162: 1867-1880[本文引用:1]
LiuK, Liu LL, Ren YL, Wang ZQ, Zhou KN, LiuX, WangD, ZhengM, Cheng ZJ, Lin QB, Wang JL, Wu FQ, ZhangX, Guo XP, Wang CM, Zhai HQ, JiangL, Wan JM. Dwarf and tiller-enhancing 1 regulates growth and development by influencing boron uptake in boron limited condition in rice. Plant Sci, 2015, 236: 18-28[本文引用:1]
[32]
TamuraW, KojimaS, ToyokawaA, WatanabeH, Tabuchi KM, HayakawaT, YamayaT. Disruption of a novel NADH-glutamate synthase2 gene caused marked reduction in spikelet number of rice. Plant Sci, 2011, 2: 1-9[本文引用:1]
[33]
Crawford NM, Ford BG. Molecular and developmental biology of inorganic nitrogen nutrition. The Arabidopsis Book, 2002[2016-03-07] http://www.aspb.org/publications/[本文引用:1]
[34]
吴巍, 赵军. 植物对氮素吸收利用的研究进展. 中国农学通报, 2010, 26(13): 75-78WuW, ZhaoJ. Advances on plants’ nitrogen assimilation and utilization. Chin Agric Sci Bull, 2010, 26(13): 75-78 (in Chinese with English abstract)[本文引用:1]
Blackwell RD, Murray A J S, Lea P J. The isolation and characterization of photorespiratory mutants of barley and pea. In: Biggins J eds. In Progress in Photosynthesis Research. Rhode Island : Springer Netherland s, 1987, pp 625-628[本文引用:1]
[37]
Somerville CR, Ogren WL. Inhibition of photosynthesis in Arabidopsis mutants lacking leaf glutamate synthase activity. Nature, 1980, 286: 257-259[本文引用:1]
 , 朱杉杉
, 朱杉杉









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}