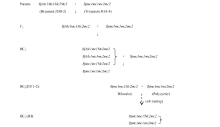
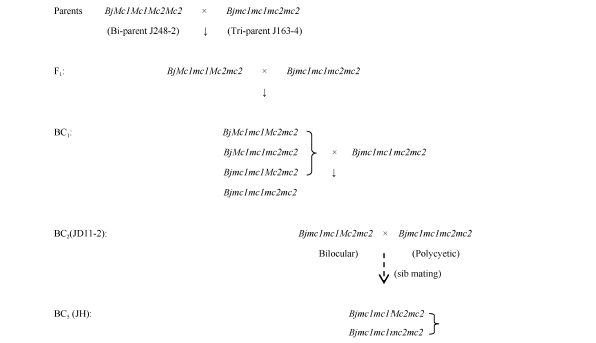
关键词:芥菜型油菜; 多室角果; BjMc2; 分子标记 Fine Mapping of Polycyetic Gene ( Bjmc2) in Brassica juncea L. WANG Gang, ZHANG Xiang-Xiang, XU Ping, LYU Ze-Wen, WEN Jing, YI Bin, MA Chao-Zhi, TU Jin-Xing, FU Ting-Dong, SHEN Jin-Xiong* National Key Laboratory of Crop Genetic Improvement / National Research Center of Rapeseed Engineering and Technology, Huazhong Agricultural University, Wuhan 430070, China Fund:This study was supported by the National Science Foundation of China (31571698). AbstractMultilocular plants have a higher seed yield than bilocular plants in B. juncea, mapping and cloning the multilocular gene(s) might be helpful for rapeseed genetic improvement and explanation of the multilocular trait development mechanism. This study verified backcross populations derived from JD11-2 family different at BjMc2 locus only can be used for BjMc2 mapping. Using AFLP assay and BSA method in BC5 and BC6 generations, one AFLP marker linked to the target gene was obtained and converted to SCAR marker (SC1). On the basis of homologous sequences of the AFLP maker in B. rapa, 11 SSR markers and one SCAR marker were identified. Through the screening in the ZBjH BAC library of Brassica juncea, two BACs covered the target area were selected and one SSR marker was developed. All the developed SCAR and SSR markers were then used to detect the BC7 population, and a linkage map for the bilocular gene BjMc2 was built. ZX17 and BACsr96, the closest flanking markers, were mapped at 0.048 cM and 0.340 cM distant from the BjMc2 gene, respectively. Bjmc2 is positioned of 68 kb between 946 kb and 1014 kb in the Scaffold 000019 physical map of A7 in B. rapa. This result would lay a foundation for cloning polycyetic gene Bjmc2and selecting polycyetic lines by marker-assisted selection.
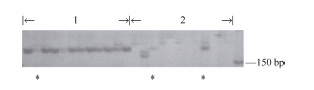
图2 与BjMc1连锁标记SR02在不同基因型单株中的扩增1~2:BjMc2位点定位群体中两室单株和多室单株; 3~4:BjMc1位点定位群体中两室单株和多室单株。Fig. 2 Amplification in diverse genotypes using marker SR02 linked toBjMc11-2: bilocular and polycyetic plants in population with BjMc2; 3-4: bilocular and polycyetic plants in population with BjMc1.
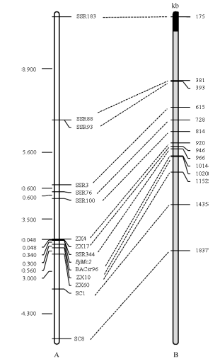
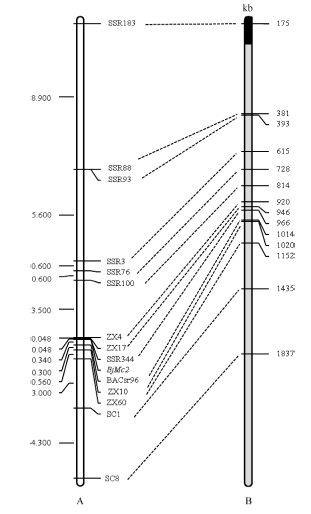
图5 芥菜型油菜两室基因BjMc2的遗传连锁图和白菜A7同源Scaffold物理图谱A是BjMc2的遗传连锁图; B是白菜A7同源Scaffold物理图谱, 黑色代表Scaffold 000101, 灰色代表Scaffold 000019。Fig. 5 Genetic linkage map of bilocular gene BjMc2 inB .junceand physical map of homologous Scaffolds of A7 in B. rapaA is the genetic linkage map of the bilocular gene BjMc2; B is physical map of homologous Scaffolds of A7 in B. rapa, black represent Scaffold 000101, gray represent Scaffold 000019.
Zhu LX, Zhang DX, Fu TD, Shen JX, WenJ. Analysis of yield and disease resistance traits of new winter rapeseed varieties over the past twenty years in China. Agric Sci Tech, 2011, 12: 842-846[本文引用:1]
[3]
Katiyar RK, ChamolaR, Chopra VL. Tetralocular mustard, Brassica juncea: new promising variability through interspecific hybridization. Plant Breed, 1998, 117: 398-399[本文引用:2]
何余堂, 龙卫华, 胡进平, 傅廷栋, 李殿荣, 陈宝元, 涂金星. 白菜型油菜角果多室性状的遗传及解剖学研究. 中国油料作物学报, 2003, 25(1): 1-4He YT, Long WH, Hu JP, Fu TD, Li DR, Chen BY, Tu JX. Anatomic and genetic studies on multicapsular character in Brassica campestris L. Chin J Oil Crop Sci, 2003, 25(1): 1-4 (in Chinese with English abstract)[本文引用:3]
吕泽文, 徐平, 张向向, 文静, 易斌, 马朝芝, 涂金星, 傅廷栋, 沈金雄. 芥菜型油菜多室角果的解剖特征及遗传分析. 中国油料作物学报, 2012, 34: 461-466Lyu ZW, XuP, Zhang XX, WenJ, YiB, Ma CZ, Tu JX, Fu TD, Shen JX. Primary study on anatomic and genetic characteristics of multi-loculus in Brassica juncea L. Chin J Oil Crop Sci, 2012, 34: 461-466 (in Chinese with English abstract)[本文引用:3]
[8]
ParitoshK, Yadava SK, GuptaV, Panjabi MP, Sodhi YS, Pradhan AK, PentalD. RNA-seq based SNPs in some agronomically important oleiferous lines of Brassica rapa and their use for genome-wide linkage mapping and specific-region fine mapping. BMC Genom, 2013, 14(1): 1-13[本文引用:3]
[9]
Fan CC, Wu YD, Yang QY, YangY, Meng QW, Zhang KQ, Li JG, Wang JF, Zhou YM. A novel single-nucleotide mutation in a CLAVATA3 gene homolog controls a multilocular silique trait in Brassica rapa L. Mol Plant, 2014, 7: 1788-1792[本文引用:3]
[10]
XiaoL, Zhao HY, ZhaoZ, Du DZ, XuL, Yao YM, Zhao ZG, Xing XR, Shang GX, Zhao HC. Genetic and physical fine mapping of a multilocular gene Bjln1 in Brassica juncea to a 208-kb region. Mol Breed, 2013, 32: 373-383[本文引用:4]
[11]
吕泽文. 芥菜型油菜多室性状的遗传分析及其基因的分子标记. 华中农业大学硕士学位论文, 湖北武汉, 2011Lyu ZW. Study on Inheritance and Molecular Markers for Multilocular Trait in Brassica junceaL. MS Thesis of Huazhong Agricultural University, Wuhan, China, 2011 (in Chinese with English abstract)[本文引用:2]
[12]
XuP, Lv ZW, Zhang XX, Wang XH, Pu YY, Wang HM, YiB, WenJ, Ma CZ, Tu JX, Fu TD. Identification of molecular markers linked to trilocular gene ( mc1) in Brassica juncea L. Mol Breed, 2014, 33: 425-434[本文引用:5]
[13]
Doyle JJ, Doyle JL. Isolation of plant DNA from fresh tissue. Focus, 1990, 12: 13-15[本文引用:1]
[14]
Michelmore RW, ParanI, Kesseli RV. Identification of markers linked to disease-resistance genes by bulked segregant analysis: a rapid method to detect markers in specific genomic regions by using segregating populations. Proc Natl Acad Sci USA, 1991, 88: 9828-9832[本文引用:1]
[15]
Negi MS, DevicM, DelsenyM, LakshmikumaranM. Identification of AFLP fragments linked to seed coat colour in Brassica juncea and conversion to a SCAR marker for rapid selection. Theor Appl Genet, 2000, 101: 146-152[本文引用:1]
刘显军. 芥菜型油菜黄籽基因克隆和黄籽形成机制分析. 湖南农业大学博士学位论文, 湖南长沙, 2013Liu XJ. Positional Cloning of The Gene for Seed Color and Molecular Mechanism of Yellow Seed Formation in Brassica juncea L. PhD Dissertation of Hunan Agricultural University, Changsha, China, 2013 (in Chinese with English abstract)[本文引用:1]
[18]
Liu RH, Meng JL. MapDraw: a microsoft excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas, 2003, 25: 317-321[本文引用:1]
Truco MJ, HuJ, SadowskiJ, Quiros CF. Inter- and intra- genomic homology of the Brassica genomes: implications for their origin and evolution. Theor Appl Genet, 1996, 93: 1225-1233[本文引用:1]
[22]
ChalhoubB, DenoeudF, Liu SY, Parkin I A P, Tang H B, Wang X Y, Chiquet J, Belcram H, Tong C B, Samans B Correa M, Da Silva C, Just J, Falentin C, Koh C S, Le Clainche I, Bernard M, Bento P, Noel B, Labadie K, Alberti A, Charles M, Arnaud D, Guo H, Daviaud C, Alamery S, Jabbari K, Zhao M, Edger P P, Chelaifa H, Tack D, Lassalle G, Mestiri I, Schnel N, Le Paslier M C, Fan G Y, Renault V, Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H D, Chalabi S, Hu Q, Fan C C, Tollenaere R, Lu Y, Battail C, Shen J S, Sidebottom C H D, Wang X F, Canaguier A, Chauveau A, Berard A, Deniot G, Guan M, Liu Z S, Sun F M, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X W, Meng J L, Ma J X, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y M, Hua W, Sharpe A G, Paterson A H, Guan C Y, Wincker P. Early allopolyploid evolution in the post-Neolithic Brassica napus oilseed genome. Science, 2014, 345: 950-953[本文引用:1]
[23]
Clark SE, Running MP, Meyerowitz EM. CLAVATA1, a regulator of meristem and flower development in Arabidopsis. Development, 1993, 119: 397-418[本文引用:3]
[24]
Clark SE, Williams RW, Meyerowitz EM. The CLAVATA1 gene encodes a putative receptor kinase that controls shoot and floral meristem size in Arabidopsis. Cell, 1997, 89: 575-585[本文引用:3]
[25]
DievartA, DalalM, Tax FE, Lacey A D. Huttly A, Li JM, Clark SE. CLAVATA1 dominant-negative alleles reveal functional overlap between multiple receptor kinases that regulate meristem and organ development. Plant Cell, 2003, 15: 1198-1211[本文引用:1]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}