关键词:小麦; 纹枯病; 蛋白激酶; TaPK-R1; 冻害 TaPK-R1Overexpressing Transgenic Wheat Lines Enhance Resistance to Sharp Eyespot and Freezing Stress LUO Mei-Ying1,2, RONG Wei2, WEI Xue-Ning2, YANG Kun2, XU Hui-Jun2, XUAN Wei-Yan1,*, ZHANG Zeng-Yan2,* 1 Agricultural College of Guangxi University, Nanning 530004, China
2 The National Key Facility for Crop Gene Resources and Genetic Improvement / Key Laboratory of Biology and Genetic Improvement of Triticeae Crops, Ministry of Agriculture / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
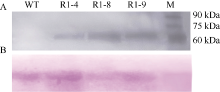
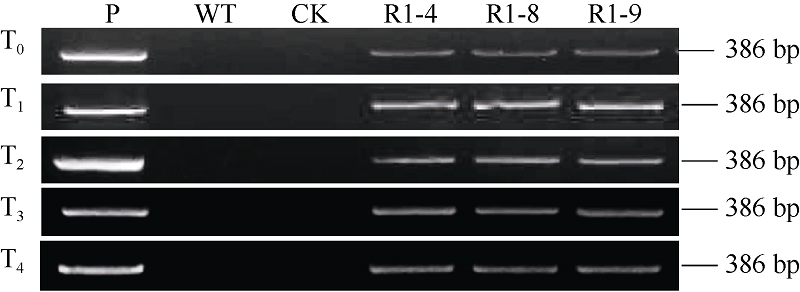
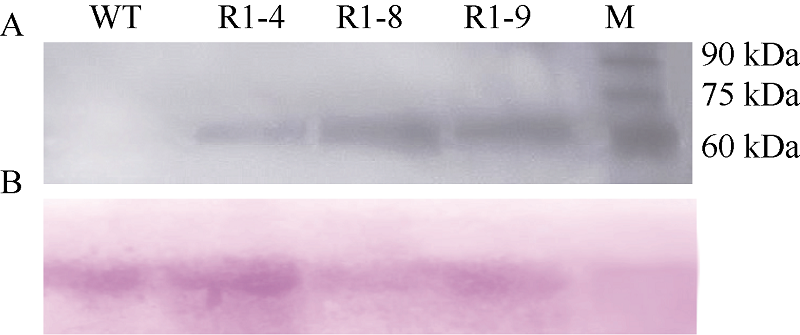
Fund:This study was supported by the National Natural Science Foundation of China (31271799) AbstractSharp eyespot caused by Rhizoctonia cerealis is a soil-borne disease and freezing stress is one of the major abiotic stresses in wheat production. The object of this study was to improve wheat resistance to sharp eyespot and freezing using transgenic technique. The transformation vector pAHC25-MYC-TaPK-R1 expressing the wheat AGC protein kinase gene TaPK-R1was constructed and transformed into the spring wheat Chinese cultivar Yangmai 20 through particle bombardment. The transformed T1 to T4 plants were subjected to PCR, RT-PCR, qRT-PCR, and Western blot analyses. Three transgenic wheat lines were generated and screened, in which the introduced TaPK-R1 transgene was inherited and expressed in higher level. After inoculation with R. cerealis virulent-isolate R0301 or WK207, these three TaPK-R1-overexpressing transgenic wheat lines displayed significant improved resistance to sharp eyespot. The infection types of these 3 transgenic lines in T1 to T4 generations were 1.16-2.11, and their disease indices were 23.20-42.10. At the same time, the infection types and disease indexes of the non-transformed wheat Yangmai 20 were 2.55-3.60 and 51.00-72.00, respectively. The three-leaf wheat seedlings were treated with -9°C for 24 hours. Freezing tolerances of the three transgenic lines were dramatically improved, whose survival rates were 52%, 79%, and 96%, respectively, and significantly higher ( P< 0.01) than that of the non-transformed Yangmai 20 (survival rate of 17%). Our results indicate that resistance/tolerance to sharp eyespot and freezing stress could be significantly enhanced in TaPK-R1 overexpressing transgenic wheat. The three transgenic lines may serve as potential resource in wheat breeding aiming at resistance improvement to sharp eyespot and freezing stress.
Keyword:Wheat; Sharp eyespot; Protein kinase; TaPK-R1; Freezing; Enhanced resistance Show Figures Show Figures
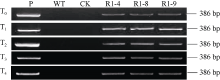
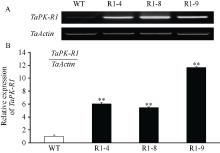
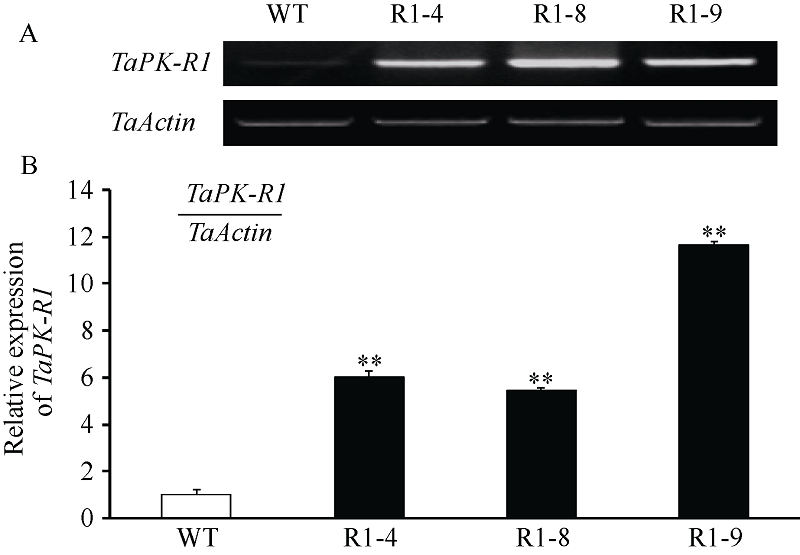
图3 转TaPK-R1基因小麦中TaPK-R1基因转录的RT-RCR(A)和qRT-PCR(B)分析 WT: 对照, 未转基因扬麦20; R1-4、R1-8和R1-9: 转TaPK-R1基因小麦材料。* * 表示转基因系与对照之间差异极显著(P< 0.01)。Fig. 3 RT-RCR (A) and qRT-PCR (B) assay on TaPK-R1 transcriptional level in the transgenic lines and its wild-type WT: wild type, untransformed wheat Yangmai 20; R1-4, R1-8, and R1-9: TaPK-R1 transgenic wheat lines. * * indicate significant difference between the transgenic line and the wild type at P < 0.01.
表1 转TaPK-R1基因小麦及受体的纹枯病抗性鉴定 Table 1 Responses ofTaPK-R1 transgenic and recipient wheat lines to R. cerealis
株系 Line
T1
T2
T3
T4
PN
IT
DI
PN
IT
DI
PN
IT
DI
PN
IT
DI
R1-4
9
1.80* *
36.0
15
1.70* *
34.0
21
1.73* *
34.6
25
1.41* *
28.2
R1-8
11
2.00* *
40.0
18
1.60* *
32.0
24
1.38* *
27.6
24
1.35* *
27.0
R1-9
12
2.11* *
42.2
17
1.48* *
29.6
21
1.16* *
23.2
24
1.30* *
26.0
WT
15
3.60
72.0
20
3.00
60.0
30
2.55
51.0
30
2.55
51.0
WT: control, untransformed wheat Yangmai 20; PN: plant number; IT: infection type; DI: disease index. The T1 and T2 plants were inoculated with R. cerealis virulent-isolate R0301, and the T3 and T4 plants were inoculated with R. cerealis virulent-isolate WK207. * * indicates significant difference between the transgenic line and the control at P< 0.01. WT: 对照, 未转基因扬麦 20; PN:植株数; IT:病级; DI: 病情指数。T1和T2代接种R. cerealis致病菌株R0301, T3和T4代接种R. cerealis致病菌株WK207。* * 表示转基因株系与对照有显著差异(P < 0.01)。
表1 转TaPK-R1基因小麦及受体的纹枯病抗性鉴定 Table 1 Responses ofTaPK-R1 transgenic and recipient wheat lines to R. cerealis
ChenL, Zhang ZY, Liang HX, Liu HX, Du LP, Xu HJ, Xin ZY. Overexpression of TiERF1 enhances resistance to sharp eyespot in transgenic wheat. J Exp Bot, 2008, 59: 4195-4204[本文引用:1]
[2]
ChenJ, Li GH, Du ZY, QuanW, Zhang HY, Che MZ, WangZ, Zhang ZJ. Mapping of QTL conferring resistance to sharp eyespot (Rhizoctonia cerealis) in bread wheat at the adult plant growth stage. Theor Appl Genet, 2013, 126: 2865-2878[本文引用:2]
[3]
梁宜策, 薛理靠, 张军峰, 王兴志. 小麦冻害调查初报. 陕西农业科学, 2003, (5): 38-41Liang YC, Xue LK, Zhang JF, Wang XZ. The investigation report of freezing injury to wheat. Shaanxi J Agric Sci, 2003, (5): 38-41 (in Chinese with English abstract)[本文引用:1]
[4]
ŁaźniewskaJ, Macioszek VK, Lawrence CB, Kononowicz AK. Fight to death: Arabidopsis thaliana defense response to fungal necrotrophic pathogens. Acta Physiol Plant, 2010, 32: 1-10[本文引用:1]
[5]
Rudd JJ, KeonJ, Hammond-Kosack K E. The wheat mitogen-activated protein kinases TaMPK3 and TaMPK6 are differentially regulated at multiple levels during compatible disease interactions with Mycosphaerella graminicola. Plant Physiol, 2008, 147: 802-815[本文引用:1]
[6]
Pearce LR, Komand erD, Alessi DR. The nuts and bolts of AGC protein kinases. Nat Rev Mol Cell Biol, 2010, 11: 9-22[本文引用:1]
[7]
DingZ, Galvan-Ampudia C S, Demarsy E, Langowski L, Kleine-Vehn J, Fan Y, Morita M T, Tasaka M, Fankhauser C, Offringa R, Friml J. Light-mediated polarization of the PIN3 auxin transporter for the phototropic response in Arabidopsis. Nat Cell BIol, 2011, 13: 447-452[本文引用:1]
[8]
Robert HS, OffringaR. Regulation of auxin transport polarity by AGC kinases. Curr Opin Plant Biol, 2008, 11: 495-502[本文引用:1]
[9]
Garcia AV, Al-YousifM, HirtH. Role of AGC kinases in plant growth and stress responses. Cellul Mol Life Sci, 2012, 69: 3259-3267[本文引用:1]
[10]
Rentel MC, LecourieuxD, OuakedF, Usher SL, PetersenL, OkamotoH, KnightH, Peck SC, Grierson CS, HirtH, Knight MR. OXI1 kinase is necessary for oxidative burst-mediated signalling in Arabidopsis. Nature, 2004, 427: 858-861[本文引用:1]
[11]
Petersen LN, Ingle RA, Knight MR, Denby KJ. OXI1 protein kinase is required for plant immunity against Pseudomonas syringae in Arabidopsis. J Exp Bot, 2009, 60: 3727-3735[本文引用:1]
[12]
Zhu XL, KunY, Wei XN, Zhang QF, WeiR, Du LP, Ye XG, QiL, Zhang ZY. The wheat AGC kinase TaAGC1 is a positive contributor to host resistance to the necrotrophic pathogen Rhizoctonia cerealis. J Exp Bot, 2015, 66: 6591-6603[本文引用:6]
[13]
Zhu XL, QiL, LiuX, Cai SB, Xu HJ, Huang RF, Li JR, Wei XN, Zhang ZY. The wheat ERF transcription factor TaPIE1 mediates host responses to both the necrotrophic pathogen Rhizoctonia cerealis and freezing stresses. Plant Physiol, 2014, 164: 1499-1514[本文引用:3]
[14]
Alexand ra SD, Konstantin VK, Valeriya SK, Olga AA. VaCPK20, a calcium-dependent protein kinase gene of wild grape vine Vitis amurensis Rupr, mediates cold and drought stress tolerance. J Plant Physiol, 2015, 185: 1-12[本文引用:1]
[15]
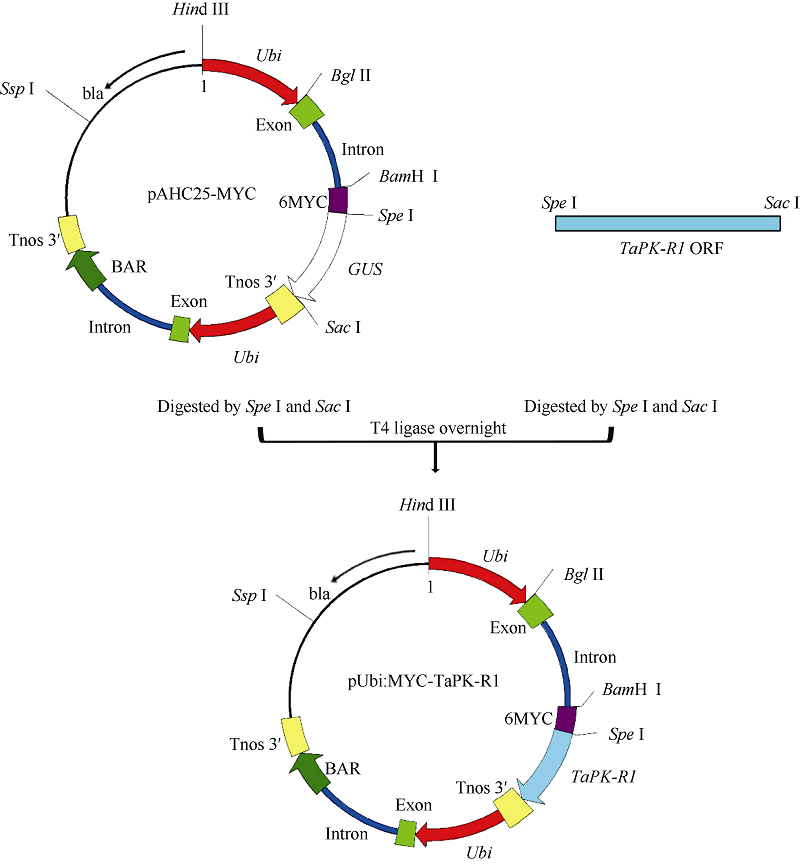
Christensen AH, Quail PH. Ubiquitin promoter-based vectors for high-level expression of selectable and /or screenable marker genes in monocotyledonous plants. Transgenic Res, 1996, 5: 213-218[本文引用:2]
[16]
徐惠君, 庞俊兰, 叶兴国, 杜丽璞, 李连城, 辛志勇, 马有志, 陈剑平, 陈炯, 程顺和, 吴宏亚. 基因枪介导法向小麦导入黄花叶病毒复制酶基因的研究. 作物学报, 2001, 27: 688-693Xu HJ, Pang JL, Ye XG, Du LP, Li LC, Xin ZY, Ma YZ, Chen JP, ChenJ, Cheng SH, Wu HY. Study on the gene transferring of Nib8 into wheat for its resistance to the Yellow mosaic virus by bombardment. Acta Agron Sin, 2001, 27: 688-694 (in Chinese with English abstract)[本文引用:1]
[17]
Saghai-Maroof MA, Soliman KM, Jorgensen RA, Allard RW. Ribosomal DNA spacer-length polymorphisms in barley: Mendelian inheritance, chromosomal location, and population dynamics. Proc Natl Acad Sci USA, 1984, 81: 8014-8019[本文引用:1]
[18]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCT method. Methods, 2001, 25: 402-408[本文引用:1]
[19]
申芳嫡, 洪彦涛, 杜丽璞, 徐惠君, 马翎健, 张增艳. 转细胞凋亡抑制基因OpIAP和p35增强小麦对纹枯病的抗性. 作物学报, 2015, 41: 1490-1499Shen FD, Hong YT, Du LP, Xu HJ, Ma LJ, Zhang ZY. Expression of Apoptosis Inhibitor genes OpIAP and p35 enhances resistance to Rhizoctonia cerealis in transgenic wheat. Acta Agron Sin, 2015, 41: 1490-1499 (in Chinese with English abstract)[本文引用:1]
[20]
魏学宁, 任丽娟, 张淼, 张巧凤, 刘欣, 周淼平, 马鸿翔, 吴继中, 马翎健, 张增艳. 人工合成抗纹枯病小麦新种植的鉴定. 植物遗传资源学报, 2015, 16: 373-378Wei XN, Ren LJ, ZhangM, Zhang QF, LiuX, Zhou MP, Ma HX, Wu JZ, Ma LJ, Zhang ZY. Identification of synthetic wheat accession with resistance to wheat sharp eyespot. J Plant Genet Resour, 2015, 16: 373-378 (in Chinese with English abstract)[本文引用:1]
[21]
王裕中, 吴志凤, 史建荣, 陈怀谷, 邵伯坤. 小麦纹枯病流行规律研究. 江苏农业科学, 1993, (麦类纹枯病专辑): 48-53Wang YZ, Wu ZF, Shi JR, Chen HG, Shao BK. Epidemic law research of wheat sharp eyespot. Jiangsu Agric Sci, 1993, (special issue of wheat sharp eyesport): 48-53 (in Chinese with English abstract)[本文引用:1]
[22]
蔡士宾, 任丽娟, 颜伟, 吴纪中, 陈怀谷, 吴小有, 张仙义. 小麦抗纹枯病种质创新及QTL定位的初步研究. 中国农业科学, 2006, 39: 928-934Cai SB, Ren LJ, YanW, Wu JZ, Chen HG, Wu XY, Zhang XY. Germplasm development and mapping of resistance to sharp eyespot (Rhizoctonia cerealis) in wheat. Sci Agric Sin, 2006, 39: 928-934 (in Chinese with English abstract)[本文引用:1]
[23]
刘颖, 张巧凤, 付必胜, 蔡士宾, 蒋彦婕, 张志良, 邓渊钰, 吴纪中, 戴廷波. 小麦纹枯病抗源的遗传多样性及抗性基因位点SSR标记分析. 作物学报, 2015, 41: 1671-1681LiuY, Zhang QF, Fu BS, Cai SB, Jiang YJ, Zhang ZL, Deng YY, Wu JZ, Dai TB. Genetic diversity of wheat germplasm resistant to sharp eyespot and genotyping of resistance loci using SSR markers. Acta Agron Sin, 2015, 41: 1671-1681 (in Chinese with English abstract)[本文引用:1]
[24]
Liu BY, LuY, Xin ZY, Zhang ZY. Identification and antifungal assay of a wheat b-1, 3-glucanase. Biotechnol Lett, 2009, 31: 1005-1010[本文引用:1]
[25]
周淼平, 周小青, 张增艳, 董娜, 马鸿翔, 姚金保. TaPIMP1过量表达提高转基因小麦纹枯病抗性的研究. 核农学报, 2011, 25: 421-426Zhou MP, Zhou XQ, Zhang ZY, DongN, Ma HX, Yao JB. Over-expression of TaPIMP1 enhanced resistance to wheat sharp eyespot in transgenic wheat. J Nucl Agric Sci, 2011, 25: 421-426 (in Chinese with English abstract)[本文引用:1]
[26]
Wei XN, Shen FD, Hong YT, RongW, Du LP, LiuX, Xu HJ, Ma LJ, Zhang Z Y. The wheat calcium-dependent protein kinase TaCPK7-D positively regulates host resistance to sharp eyespot disease. Mol Plant Pathogen, 2016, online. doi: DOI:10.1111/mpp12360[本文引用:1]
[27]
Liu ST, Liu SW, WangM, Wei TT, MengC, WangM, Xia GM. A wheat similar to RCD-ONE gene enhances seedling growth and abiotic stress resistance by modulating redox homeostasis and maintaining genomic integrity. Plant Cell, 2014, 26: 164-180[本文引用:1]
[28]
XiangY, HuangY, XiongL. Characterization of stress-responsive CIPK genes in rice for stress tolerance improvement. Plant Physiol, 2007, 44: 1416-1428[本文引用:2]
[29]
CaiG, WangG, WangL, PanJ, LiuY, LiD. ZmMKK1, a novel group A mitogen-activated protein kinase gene in maize, conferred chilling stress tolerance and was involved in pathogen defense in transgenic tobacco. Plant Sci, 2014, 214: 57-73[本文引用:1]
 , 荣玮
, 荣玮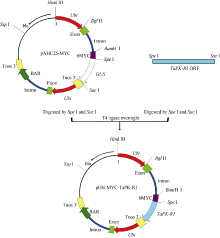



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}