关键词:小麦DELLA蛋白; NaCl胁迫; 叶绿素含量; 超氧化物歧化酶; 丙二醛 A Wheat DELLA Gain-of-function Mutant Aibian 1 Promotes Seedling Salt Tolerance WANG Run-Qing1,2,**, FAN Xiao-Cong2,3,**, SONG Mei-Fang2,4, XIAO Yang5, GUO Lin2, MENG Fan-Hua2, YANG Qing-Hua3, WU Da-Fu1,*, YANG Jian-Ping1,2,* 1School of Life Science & Technology, Henan Institute of Science and Technology, Xinxiang 453000, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
3College of Agronomy, Henan Agricultural University / Collaborative Innovation Center of Henan Grain Crops, Zhengzhou 450002, China
4 Beijing Radiation Center, Beijing 100875, China
5 Graduate School of Chinese Academy of Agricultural Sciences, Beijing 100081, China
Fund:This study was supported by the National Key R&D Program of China (Pilot Project on Seven Main Crop Breeding) (2016YFD101802 ), the Natural Science Foundation of Beijing (Key Program) (6151002) and the Agricultural Science and Technology Innovation Program of CAAS AbstractDELLA proteins are negative regulators in GA signal pathway. DELLA protein is able to response to various environmental signals and plays an important role in plant resistance to biotic and abiotic stresses. The DELLA gain-of-function mutant Aibian 1 has been widely applied in wheat breeding for dwarfness, but its tolerance to salt is unclear. In this study, Aibian 1 was compared with Chinese Spring, Jing 411, and Aiganzao in salt tolerance using Hoagland’s hydroponic culture with 200 mmol L-1 NaCl. Total chlorophyll content, relative water content, superoxide dismutase (SOD) activity and malondialdehyde (MDA) content were measured to evaluate the ability of salt tolerance. Immunoblot assay was employed to assess the DELLA protein abundance in wheat seedlings. After seven-day NaCl treatment, Aibian 1 showed no clear withering leaves, whereas the other three cultivars displayed obviously wilting and yellowing leaves. Among the four cultivars, Aibian 1 had the least loss on chlorophyll content and relative water content, the highest SOD activity, and the lowest levels of MDA content after salt treatment. Immunoblot assay indicated the highest accumulation of DELLA protein in Aibian 1 among the four cultivars. Thus, we consider the high level of DELLA protein is closely related to salt tolerance in Aibian 1.
Keyword:Wheat DELLA proteins; Salt tolerance; Chlorophyll content; Superoxide dismutase; Malondialdehyde Show Figures Show Figures
图1 盐胁迫前后的小麦幼苗形态 小麦种子28℃催芽24 h后在蛭石中22℃生长4 d (A和B), 然后在Hoagland液(pH 6.0)中缓冲2 d, 再分别转入不含(C)或含(D) 200 mmol L-1 NaCl的Hoagland培养液(pH 6.0)生长7 d。E图是D图方框部分的局部放大。1: 中国春; 2: 京411; 3: 矮秆早; 4: 矮变1号。Bar = 2 mm。Fig. 1 Morphology of wheat seedlings in response to salt stress Wheat seeds were germinated at 28 ° C for 24 h and grown in vermiculite for 4 d in greenhouse at 22 ° C (panels A and B). Wheat seedlings were carefully transferred into the Hoagland’ s solution (pH 6.0) for 2 d to buffer and finally cultured in Hoagland’ s solution without (panel C) or with (panel D) 200 mmol L-1 NaCl for 7 d. Panel E is the enlargement of framed parts in panel D. 1: Chinese Spring; 2: Jing 411; 3: Aiganzao; 4: Aibian 1. Bar = 2 mm.
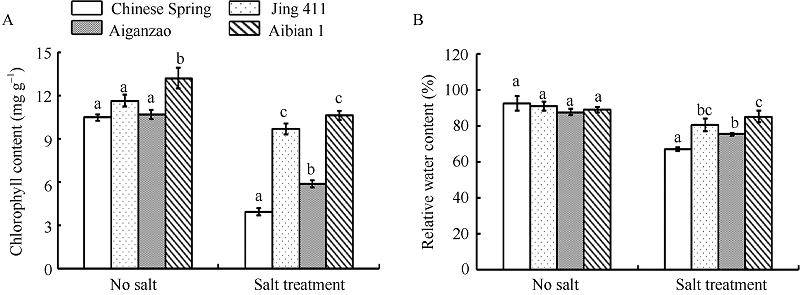
图2 盐胁迫和无胁迫下4个小麦品种的叶绿素含量(A)和相对含水量(B)变化 数据为3次生物学重复的平均值和标准差。Fig. 2 Changes of chlorophyll content (A) and relative water content (B) in four wheat cultivars under treatment with or without salt stress Data are the means and standard deviations of three biological repeats.
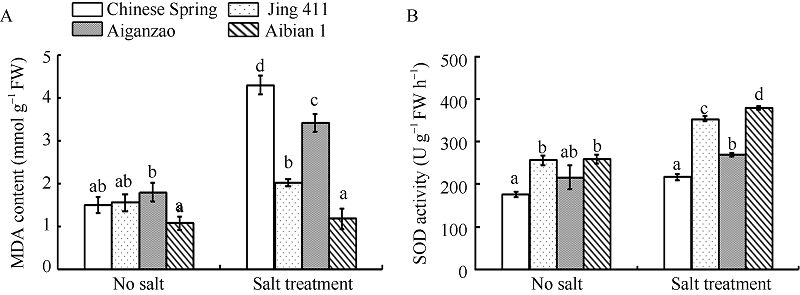
图3 盐胁迫和无胁迫下4个小麦品种中MDA含量(A)和SOD活性(B)的变化 数据为3次生物学重复的平均值和标准差。Fig. 3 Changes of MDA content (A) and SOD activity (B) in four wheat cultivars under treatments with or without salt stress Data are the means and standard deviations of three biological repeats.
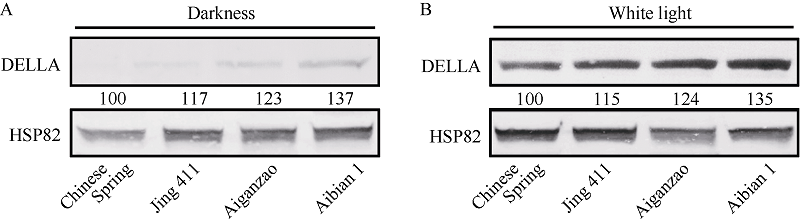
图4 4个小麦品种的幼苗在白光或黑暗下DELLA蛋白的免疫印迹检测 A: 22℃白光(30.0 µ mol m-2 s-1)生长6 d; B: 22℃黑暗生长6 d。HSP82免疫印迹显示大约相当的上样量。Fig. 4 Immunoblot analysis of DELLA protein accumulation in four wheat cultivars under white light or darkness A: Seedlings were grown for six days under 22° C and white light (30.0 µ mol m-2 s-1); B: Seedlings were grown for six days under 22℃ and darkness. Anti-HSP82 immunoblots indicates approximately equal loading.
王佳丽, 黄贤金, 钟太洋, 陈志刚. 盐碱地可持续利用研究综述. 地理学报, 2011, 66: 673-684Wang JL, Huang XJ, Zhong TY, Chen ZG. Review on sustainable utilization of salt-affected land . Acta Geograph Sin, 2011, 66: 673-684 (in Chinese with English abstract)[本文引用:1]
[2]
嵇怡, 缪旻珉, 陈学好. 植物矮生性状的分子遗传研究进展. 分子植物育种, 2007, 4: 753-771JiY, Miao MM, Chen XH. Progresses on the molecular genetics of dwarf character in plants. Mol Plant Breed, 2006, 4: 753-771 (in Chinese with English abstract)[本文引用:1]
[3]
Li YY, Xiao JH, Wu JJ, Duan JL, LiuY, Ye XG, ZhangX, Guo XP, Gu YQ, Zhang LC, Jia JZ, Kong XY. A tand em segmental duplication (TSD) in green revolution gene Rht-D1b region underlies plant height variation. New Phytol, 2012, 196: 282-291[本文引用:3]
[4]
BarbaraF, Harberd NP. Evidence that the Arabidopsis nuclear gibberellin signalling protein GAI is not destabilised by gibberellin. Plant J, 2003, 32: 935-947[本文引用:2]
[5]
Pysh LD, Wysocka-Diller J W, Camilleri C, The GRAS gene family in Arabidopsis: sequence characterization and basic expression analysis of the SCARECROW-LIKE genes. Plant J, 1999, 18: 111-119[本文引用:1]
[6]
ItonH, Ueguchi-TanakaM, SatoY, AshikariM, MatsuokaM. The gibberellin signaling pathway is regulated by the appearance and disappearance of SLENDER RICE1 in nuclei. Plant Cell, 2002, 14: 57-70[本文引用:1]
[7]
Wen CK, ChangC. Arabidopsis RGL1 encodes a negative regulator of gibberellin responses. Plant Cell, 2002, 14: 87-100[本文引用:1]
[8]
FuX, Harberd NP. Auxin promotes Arabidopsis root growth by modulating gibberellin response. Nature, 2003, 421: 740-743[本文引用:1]
[9]
CaoD, HussainA, ChengH. Loss of function of four DELLA genes leads to light and gibberellin-independent seed germination in Arabidopsis. Planta, 2005, 223: 105-113[本文引用:1]
[10]
PengJ, Richards DE, Hartley NM, Murphy GP, Devos KM, Flintham JE, BealesJ, Fish LJ, Worland AJ, PelicaF, SudhakarD, ChristouP, Snape JW, Gale MD, Harberd NP. “Green revolution” genes encode mutant gibberellin response modulators. Nature, 1999, 400: 256-261[本文引用:2]
[11]
Boss PK, Thomas MR. Association of dwarfism and floral induction with a grape ‘green revolution’ mutation. Nature, 2002, 416: 847-850[本文引用:1]
[12]
MagomeH, YamaguchiS, HanadaA, KamiyaY, OdaK. The DDF1 transcriptional activator upregulates expression of a gibberellin-deactivating gene, GA2ox7, under high-salinity stress in Arabidopsis. Plant J, 2008, 56: 613-626[本文引用:1]
MagomeH, YamaguchiS, HanadaA, KamiyaY, OdaK. Dwarf and delayed-flowering 1, a novel Arabidopsis mutant deficient in gibberellin biosynthesis because of overexpression of a putative AP2 transcription factor. Plant J, 2004, 37: 720-729[本文引用:1]
[15]
AchardP, GongF, CheminantS, AliouaMalek, HeddenP, GenschikaP. The cold-inducible CBF1 factor-dependent signaling pathway modulates the accumulation of the growth-repressing DELLA proteins via its effect on gibberellin metabolism. Plant Cell, 2008, 20: 2117-2129[本文引用:1]
[16]
JiangC, GaoX, LiaoLL, Harberd NP, Fu XD. Phosphate starvation root architecture and anthocyanin accumulation responses are modulated by the gibberellin-DELLA signaling pathway in Arabidopsis. Plant Physiol, 2007, 145: 1460-1470[本文引用:1]
[17]
黄先忠, 马正强. DELLA家族蛋白与植物生长发育的关系. 植物生理通讯, 2004, 40: 529-532Huang XZ, Ma ZQ. Progress in studies on DELLA protein family in plant growth and development. Plant Physiol Commun, 2004, 40: 529-532 (in Chinese)[本文引用:1]
[18]
陆维忠, 赵寅槐, 冯晓棠, 邹明烈. 小麦“矮变1号”矮秆遗传的初步分析. 作物学报, 1982, 8: 65-66Lu WZ, Zhao YH, Feng XT, Zou ML. A preliminary study on the inheritance of dwarfness of wheat variety Aibian 1. Acta Agron Sin, 1982, 8: 65-66 (in Chinese)[本文引用:1]
[19]
范瑞英. 矮败小麦在西藏的进展及应用研究. 西藏农业科技, 2008, 30(4): 15-17Fan RY. The study progress and application of dwarf male-sterile wheat in Tibet. Tibetan J Agric Sci, 2008, 30(4): 15-17 (in Chinese)[本文引用:1]
[20]
孙广华, 原换换, 樊小聪, 顾海科, 宋梅芳, 肖阳, 孟凡华, 郭林, 杨青华, 詹克慧, 杨建平. 甘蓝光敏色素B基因的克隆及在拟南芥中异源转基因的功能验证. 中国农业科学, 2015, 48: 4417-4427Sun GH, Yuan HH, Fan XC, Gu HK, Song MF, XiaoY, Meng FH, GuoL, Yang QH, Zhan KH, Yang JP. Molecular cloning and Arabidopsis ectopic expression of a phytochrome B gene from Brassica oleracea. Sci Agric Sin, 2015, 48: 4417-4427 (in Chinese with English abstract)[本文引用:2]
[21]
万里强, 石永红, 李向林, 何峰, 贾亚雄. PEG 胁迫下3个多年生黑麦草品种抗性生理研究, 草地学报, 2009, 17: 440-444Wan LQ, Shi YH, Li XL, HeF, Jia YX. Physiological resistance of three Lolium perenne L. varieties under PEG stress. Acta Agrest Sin, 2009, 17: 440-444 (in Chinese with English abstract)[本文引用:1]
[22]
孙群, 胡景江. 植物生理学研究技术. 陕西杨凌: 西北农林科技大学出版社, 2006. pp 165-177SunQ, Hu JJ. Technologies for Plant Physiology Reasearch, Yangling: Northwest Agriculture and Forestry University of Science and Technology Press, 2006. pp 165-177(in Chinese)[本文引用:2]
[23]
ZhengX, Wu SW, Zhai HQ, ZhouP, Song MF, SuL, Xi YL, Li ZY, Cai YF, Meng FH, YangL, Wang HY, Yang JP. Arabidopsis phytochrome B promotes SPA1 nuclear accumulation to repress photomorphogenesis under far-red light. Plant Cell, 2013, 25: 115-133[本文引用:1]
李彦, 张英鹏, 孙明, 高弼模. 盐分胁迫对植物的影响及植物耐盐机理研究进展. 中国农学通报, 2008, 24(1): 258-265LiY, Zhang YP, SunM, Gao BM. Research advance in the effects of salt stress on plant and the mechanism of plant resistance. Chin Agric Sci Bull, 2008, 24(1): 258-265 (in Chinese with English abstract)[本文引用:1]
[26]
华春, 王仁雷. 盐胁迫对水稻叶片光合效率和叶绿体超显微结构的影响. 山东农业大学学报(自然科学版), 2004, 35(1): 27-31HuaC, Wang RL. Salt stress affects photosynthetic efficiency and chloroplast ultrastructure of rice leaves. J Shand ong Agric University (Nat Sci Edn), 2004, 35(1): 27-31 (in Chinese with English abstract)[本文引用:1]
[27]
Lee DH, Kim YS, Lee CB. The inductive responses of the antioxidant enzymes by salt stress in the rice (Oryza sativa L. ). J Plant Physiol, 2001, 158: 737-745[本文引用:1]
[28]
Hernand ez JA, JimenezA, MullineauxP, SevillaF. Tolerance of pea (Pisum sativum L. ) to long-term salt stress is associated with induction of antioxidant defences. Plant Cell Environ, 2000, 23: 853-862[本文引用:1]
[29]
熊国胜, 李家洋, 王永红. 植物激素调控研究进展. 科学通报, 2009, 54: 2718-2733Xiong GS, Li JY, Wang YH. Research progress of plant hormone regulation. Chin Sci Bull, 2009, 54: 2718-2733 (in Chinese)[本文引用:1]
[30]
SchwechheimerC. Understand ing gibberellic acid signaling-are we there yet?Curr Opin Plant Biol, 2008, 11: 9-15[本文引用:1]
[31]
AchardP, Renou JP, BerthoméR. Plant DELLAs restrain growth and promote survival of adversity by reducing the levels of reactive oxygen species. Curr Biol, 2008, 18: 656-660[本文引用:1]
 , 樊晓聪
, 樊晓聪





{kind=link}
{kind=link}
{kind=link}
{kind=link}