关键词:水稻; 稻瘟病; 抗病基因; 极端分离群体分池法; 抗性基因聚合 Analysis of Blast Resistance Genes in Japonica Variety Jijing 809 ZHU Ya-Jun1,**, SUN Qiang2,**, WANG Jin-Ming2, CHEN Kai1, FENG Bo1, FANG Ya-Jie1, LIN Xiu-Yun2,*, XU Jian-Long1,3,* 1 National Facility for Crop Gene Resources and Genetic Improvement / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2 Rice Research Institute, Jilin Academy of agricultural Sciences, Gongzhuling 130000, China
3 Shenzhen Institute of Breeding and Innovation, Chinese Academy of Agricultural Sciences, Shenzhen 518120, China
Fund:This study was supported by the Program of Introducing International Super Agricultural Science and Technology (2010-G2B-2), Project of transformation of scientific and technological achievements in Jilin Province (20130305038NY), the National High Technology Research and Development Program of China (2014AA10A601), and the Natural Science Foundation of Jilin Province (20140101014JC) AbstractBlast is one of the most hazardous diseases in rice production. Planting resistance variety is an effective solution to control the disease. In this study, a backcross population derived from two parents, Jijing 88 and 93072, was selected to identify blast resistance genes using bulked extremes and recessive class under artificial inoculation and further to deduce the composition of resistance genes in Jijing 809 developed from Jijing 88 and 93072. Four major resistance genes, i.e., Pi-2(t), Pi-7-1(t), Pi-7-2(t), and Pi-11(t), were responsible for segregation of resistance to the strong virulent strain GD9-1, and only one resistance gene Pi-2(t) for segregation of resistance to the weak virulent strain GD19-1. Among them, Pi-2(t) was effective to both strains. The favorable alleles at all loci were from 93072 except for the allele at Pi-2(t) from Jijing 88. According to genomic comparison, Pi-2(t) was deduced to be allelic to Pi-b, and Pi-11(t) to Pi-47(t) or Pik; whereas Pi-7-1(t)and Pi-7-2(t) were two novel resistance genes, which were linkages to SSR markers with RM21260 (0.11 cM) and RM8037 (6.97 cM). Genotypes on the four above-referenced loci were compared between Jijing 809 and its parents by using closely linked SSR markers and a set of 56k gene chip developed from re-sequenced data of 3000 accessions of rice germplasm. The results indicated that the resistance genes in Jijing 809, Pi-2(t) and Pi-11(t), were inherited from the recurrent parent Jijing 88 and the donor parent 93072, respectively, which reasonably explained the higher blast resistance in Jijing 809 than in Jijing 88. Finally, we discussed how to pyramid different known major resistant genes, especially to make full use of the ‘defeated’ resistance gene in the original resistant variety to improve blast resistance in rice.
Keyword:Rice; Blast; Resistance gene; Bulked extremes and recessive class; Pyramiding of resistance gene Show Figures Show Figures
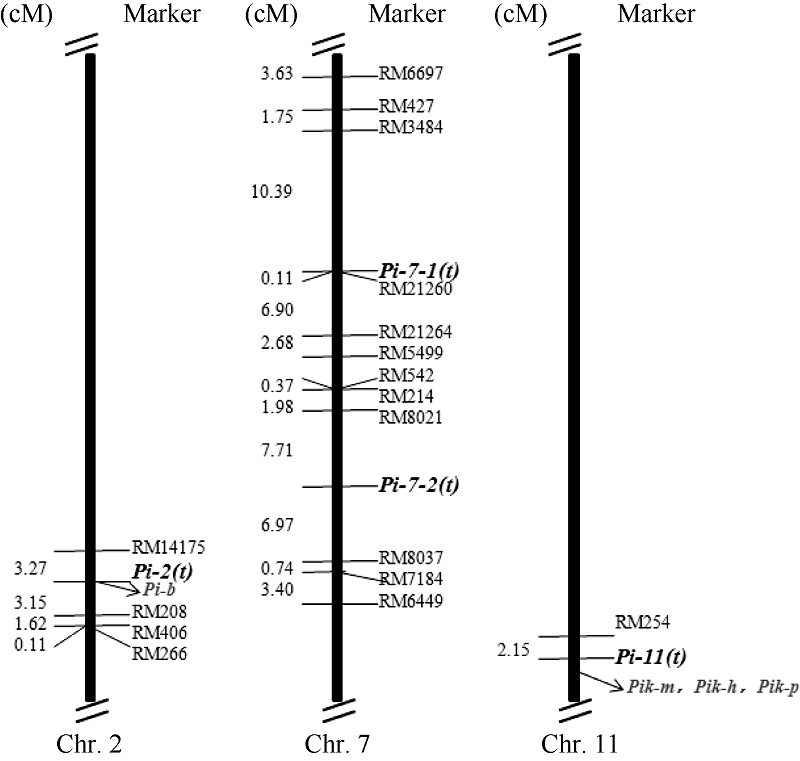
图1 连锁图谱和抗性基因在染色体上的分布 粗黑体字体为本研究中定位到的抗性基因, 灰色字体为已经克隆的抗性基因。Fig. 1 Genetic linkage map and distribution of four resistance genes on chromosomes The bold fonts indicate the rice blast resistance genes mapped in this study, and the grey ones indicate the rice blast resistance genes previously cloned.
4 结论定位到4个主效抗病基因, 其中Pi-2(t)同时抗2个菌株。Pi-2(t)可能与Pi-b等位, Pi-11(t)可能与Pi-47(t)或Pik等位, Pi-7-1(t)和Pi-7-2(t)是2个新的抗性位点, 除Pi-2(t)位点的抗性等位基因来自吉粳88外, 其余3个位点的抗病等位基因均来自93072。吉粳809携带来自受体亲本吉粳88的Pi-2(t)和来自供体亲本93072的Pi-11(t) 2个抗性基因, 合理地解释了吉粳809抗性明显好于吉粳88的原因。 致谢: 感谢广东省农业科学院植物保护研究所朱小源研究员提供苗期稻瘟病接种帮助。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Wilson RA, Talbot NJ. Under pressure: investigating the biology of plant infection by Magnaporthe oryzae. Nat Rev Microbiol, 2009, 7: 185-195[本文引用:1]
[2]
孙国昌, 杜新法, 陶荣祥, 孙淑沅. 水稻稻瘟病防治策略和21世纪研究展望. 植物病理学报, 1998, 28: 289-292Sun GC, Du XF, Tao RX, Sun SY. Control tactics and prospect of rice blast research in 21th century. Acta Phytopath Sin, 1998, 28: 289-292 (in Chinese)[本文引用:2]
[3]
MoytriR, Jia YL, Richard DC. Structure, function, and co-evolution of rice blast resistance genes. Acta Agrin Sin, 2012, 38: 381-393[本文引用:1]
[4]
全国农业技术推广服务中心. 2012年全国农作物重大病虫害呈重发态势. 农药市场信息, 2012, (5): 49The National Agro-techExtension and ServiceCenter. The trends caused by major diseases and pests in crops will be severe in 2012 in China. Pestcide Market News, 2012, (5): 49 (in Chinese)[本文引用:1]
[5]
江南, 刘雄伦, 戴良英, 王国梁. 水稻抗稻瘟病基因的定位与克隆研究进展. 中国农学通报, 2010, 26(10): 270-275JiangN, Liu XL, Dai LY, Wang GL. Advances on the mapping and cloning of blast resistance gene in rice. Chin Agric Sci Bull, 2010, 26(10): 270-275 (in Chinese with English abstract)[本文引用:1]
[6]
韦新宇, 许旭明, 张锐, 陈美莲, 马彬林, 邹文广, 杨旺兴, 卓伟, 王宗华, 梁康迳. 籼粳交新种质康丰A对稻瘟病抗性的遗传. 植物遗传资源学报, 2014, 15: 113-1137Wei XY, Xu XM, ZhangR, Chen ML, Ma BL, Zou WG, Yang WX, ZhuoW, Wang ZX, Liang KJ. Inheritance of blast resistance in new germplasm Kangfeng A from indica-japonica crosses. J Plant Genet Res, 2014, 15: 113-1137 (in Chinese with English abstract)[本文引用:1]
[7]
何秀英, 王玲, 吴伟怀, 陈钊明, 林菲, 程永盛, 刘维, 陈粤汉, 廖耀平. 水稻稻瘟病抗性基因的定位、克隆及育种应用进展. 中国农学通报, 2014, 30(6): 1-12He XY, WangL, Wu WH, Chen ZM, LinF, Cheng YS, LiuW, Chen YH, Liao YP. The progress of mapping, isolation of the genes resisting to blast and their breeding application in rice. Chin Agric Sci Bull, 2014, 30(6): 1-12 (in Chinese with English abstract)[本文引用:1]
[8]
BalliniE, Morel JB, DrocG, PriceA, CourtoisB, Notteghem JL, TharreauD. Genome-wide meta-analysis of rice blast resistance genes and quantitative trait loci provides new insights into partial and complete resistance. Mol Plant Microbe Interac, 2008, 21: 859-868[本文引用:2]
[9]
LiuY, LiuB, Zhu XY, Yang JY, BordeosA, Wang GL, LeachJ, LeungH. Fine-mapping and molecular marker development for Pi56(t), a NBS-LRR gene conferring broad-spectrum resistance to Magnaporthe oryzae in rice. Theor Appl Genet, 2013, 126: 985-998[本文引用:2]
杨勤忠, 林菲, 冯淑杰, 王玲, 潘庆华. 水稻稻瘟病抗性基因的分子定位及克隆研究进展. 中国农业科学, 2009, 42: 1601-1615Yang QZ, LinF, Feng SJ, WangL, Pan QH. Recent progress on molecular mapping and cloning of blast resistance genes in rice (Oryza sativa L. ). Sci Agric Sin, 2009, 42: 1601-1615 (in Chinese with English abstract)[本文引用:1]
[12]
Sharma TR, Rai AK, Gupta SK, VijayanJ, Devanna BN, RayS. Rice blast management through host-plant resistance: retrospect and prospects. Agric Res, 2012, 1: 37-52[本文引用:1]
[13]
张俊国, 张三元, 全成哲, 杨春刚, 郭桂珍, 孙强. 超级稻吉粳88的优异特性及推广应用. 中国稻米, 2008, 31(3): 42-44Zhang JG, Zhang SY, Quan CZ, Yang CG, Guo GZ, SunQ. The properties and application of Jijing 88, a super rice. China Rice, 2008, 31(3): 42-44 (in Chinese)[本文引用:2]
[14]
王金明, 战长春, 高明晶, 孙强, 林秀云. 水稻新品种吉粳809选育报告. 北方水稻, 2013, 43(3): 58-59Wang JM, Zhan CC, Gao MJ, SunQ, Lin XY. Breeding report on Jijing 88, a new rice variety. North Rice, 2013, 43(3): 58-59 (in Chinese)[本文引用:1]
[15]
张三元, 张俊国, 赵劲松, 李彻, 林秀云, 郭桂珍, 杨春刚, 孙强. 优质、高产、多抗水稻新品种吉粳88选育研究. 作物研究, 2007, (3): 222-226Zhang SY, Zhang JG, Zhao JS, LiC, Lin XY, Guo GZ, Yang CG, SunQ. Breeding of Jijing 88, a high quality, high yield and multi-resistance rice variety. Crop Res, 2007, (3): 222-226 (in Chinese)[本文引用:1]
[16]
杨健源, 陈深, 曾列先, 李亦龙, 陈珍, 朱小源. 稻瘟病主效抗性基因对广东省籼稻稻瘟病菌的抗性评价. 中国水稻科学, 2008, 22: 190-196Yang JY, ChenS, ZengLX, Li YL, ChenZ, Zhu XY. Evaluation on resistance of major rice blast resistance genes to Magnaporthe grisea isolates collected from indica rice in Guangdong province, china. Chin J Rice Sci, 2008, 22: 190-196 (in Chinese with English abstract)[本文引用:1]
[17]
IRRI. Stand ard Evaluation System for Rice. Manila, Philippines: IRRI, 2002[本文引用:1]
[18]
Tan CX, Ji XM, YangY, Pan XY, Zuo SM, Zhang YF, Zou JH, Chen ZX, Zhu LH, Pan XB. Identification and marker-assisted selection of two major quantitative genes controlling rice sheath blight resistance in backcross generations. Acta Genet Sin, 2005, 32: 399-405[本文引用:1]
[19]
Alexand rovN, TaiS, Wang WS, MansuetoL, PalisK, Fuentes RR, Ulat VJ, ChebotarovD, Zhang GY, Li ZK, MauleonR, Hamilton RS, McNally K L. SNP-Seek database of SNPs derived from 3000 rice genomes. Nucl Acids Res, 2015, 43: D1023-D1027[本文引用:1]
[20]
FjellstromR, Conaway-Bormans C A, McClung A M, Marchetti M A, Shank A R, Park W D. Development of DNA markers suitable for marker assisted selection of three Pi genes conferring resistance to multiple Pyricularia grisea pathotypes. Crop Sci, 2004, 44: 1790-1798[本文引用:1]
[21]
KoideY, KawasakiA, Telebanco-Yanoria M J, Hairmansis A, Nguyet N T M, Bigirimana J, Fujita D, Kobayashi N, Fukuta Y. Development of pyramided lines with two resistance genes, Pish and Pib, for blast disease (Magnaporthe oryzae B. Couch) in rice (Oryza sativa L. ). Plant Breed, 2010, 129: 670-675[本文引用:1]
[22]
沈海波, 付胜, 杜文鑫, 林秀云, 孙强, 张海波, 王金明. 吉林省主推水稻品种抗稻瘟病基因的分子检测. 安徽农业科学, 2013, 41: 9186-9189Shen HB, FuS, Du WX, Lin XY, SunQ, Zhang HB, Wang JM. Molecular detection of blast resistance genes of major varieties of rice in Jilin province. J Anhui Agric Sci, 2013, 41: 9186-9189 (in Chinese with English abstract)[本文引用:1]
[23]
Tanweer FA, Rafii MY, SijamS, Rahim HA, AhmedF, AshkanS, LatifA. Identification of suitable segregating SSR markers for blast resistance in rice using inheritance and disease reaction analysis in backcross families. Aust Plant Pathal, 2015, 44: 619-627[本文引用:1]
[24]
Pan QH, TanisakaT, IkehashiH. Studies on the genetics and breeding of blast resistance in rice: VI. Gene analysis for the blast resistance of two Yunnan native cultivars GA20 and GA25. Breed Sci, 1996, 46(S2): 70[本文引用:1]
[25]
Li LY, WangL, Jing JX, Li ZQ, LinF, Huang LF, Pan QH. The Pikm gene, conferring stable resistance to isolates of Magnaporthe oryzae, was finely mapped in a crossover-cold region on rice chromosome 11. Mol Breed, 2007, 20: 179-188[本文引用:1]
[26]
Huang HM, HuangL, Feng GP, Wang SH, WangY, Liu JL, JiangN, Yan WT, Xu LC, Sun PY, Liu ZQ, Pan SJ, Liu XL, Xiao YH, Liu EM, Dai LY, Wang GL. Molecular mapping of the new blast resistance genes Pi47 and Pi48 in the durably resistant local rice cultivar Xiangzi 3150. Phytopathology, 2011, 101: 620-626[本文引用:1]
[27]
Li ZK, Pinson S R M, Patemon A H, Parks W D, Stansel J W. Genetics of hybrid sterility and hybrid breakdown in an intersubspecific rice (Oryza sativa L. ) population. Genetics, 1997, 145: 1139-1148[本文引用:1]
[28]
ZhangF, Zhuo DL, Zhang LY, Wang WS, Xu JL, Vera-Cruz C M, Li Z K, Zhou Y L. Xa39, a novel dominant gene conferring broad-spectrum resistance to Xanthomonas oryzae pv. oryzae in rice. Plant Pathol, 2015, 64: 568-575[本文引用:1]
[29]
ServinB, Martin OC, MézardM, HospitalF. Toward a theory of marker-assisted gene pyramiding. Genetics, 2004, 168: 513-523[本文引用:1]
[30]
沈瑛, AderitH, 朱旭东, Milazzo J, 陈红旗, Tharreau D. 中国部分杂交稻和常规早籼、晚粳品种(系)的抗瘟性. 中国农业科学, 2004, 37: 362-369ShenY, AdreitH, Zhu XD, MilazzoJ, Chen HQ, TharreauD. Evaluation of resistance of some hybrid rice, conventional early indica and late japonica rice to Magnaporthe grisea in China. Sci Agric Sin, 2004, 37: 362-369 (in Chinese with English abstract)[本文引用:1]
[31]
HittalmaniS, ParcoA, MewTV, Zeigler RS, HuangH. Fine mapping and DNA marker-assisted pyramiding of the three major genes for blast resistance in rice. Theor Appl Genet, 2000, 100: 1121-1128[本文引用:1]
[32]
柳武革, 王丰, 金素娟, 朱小源, 李金华, 刘振荣, 廖亦龙, 朱满山, 黄慧君, 符福鸿, 刘宜柏. 利用分子标记辅助选择聚合Pi-1和Pi-2基因改良两系不育系稻瘟病抗性. 作物学报, 2008, 34: 1128-1136Liu WG, WangF, Jin SJ, Zhu XY, Li JH, Liu ZR, Liao YL, Zhu MS, Huang HJ, Fu FH, Liu YB. Improvement of rice blast resistance in TGMS line by pyramiding of Pi-1 and Pi-2 through molecular marker-assisted selection. Acta Agron Sin, 2008, 34: 1128-1136 (in Chinese with English abstract)[本文引用:1]
[33]
李洪亮, 李荣田. 稻瘟病抗性基因Pi1和Pi2的聚合及其育种价值分析. 北方水稻, 2010, 40(5): 7-12Li HL, Li RT. The polymerization of rice-blast resistance genes Pi1 and Pi2 and analysis on breeding value. North Rice, 2010, 40(5): 7-12 (in Chinese with Eng lish abstract)[本文引用:1]
[34]
王军, 杨杰, 陈志德, 范方军, 朱金燕, 杨金欢, 仲维功. 利用分子标记辅助选择聚合水稻抗病基因Pi-ta、Pi-b和Stv-bi. 作物学报, 2011, 37: 975-981WangJ, YangJ, Chen ZD, Fan FJ, Zhu JY, Yang JH, Zhong WG. Pyramiding resistance gene Pi-ta, Pi-b, and Stv-bi by marker-assisted selection in rice (Oryza sativa L. ). Acta Agron Sin, 2011, 37: 975-981 (in Chinese with English abstract)[本文引用:1]
[35]
向小娇, 张建, 郑天清, 徐建龙. 应用分子标记技术改良京作1号的稻瘟病抗性. 植物遗传资源学报, 2016, 17: 772-779Xiang XJ, ZhangJ, Zheng TQ, Xu JL. Improving blast resistance of Jingzuo 1 using molecular marker technique. J Plant Genet Res, 2016, 17: 772-779 (in Chinese with English abstract)[本文引用:1]
[36]
程芳艳, 李春光, 刘永巍, 孙翊轩, 孟昭河, 徐正进. 水稻抗瘟基因Pi-b与Pi-5的分子检测及外引抗源的利用评价. 核农学报, 2016, 30: 11-18Cheng YF, Li CG, Liu YW, Sun YX, Meng ZH, Xu ZJ. Molecular detection of blast resistance genes Pi-b and Pi-5 in northeast rice varieties and utilization evaluation of introduced resistant donors. Acta Agric Nucl Sin, 2016, 30: 11-18 (in Chinese English abstract)[本文引用:1]
[37]
Li ZK, Luo LJ, Mei HW, PatersonH, Zhao XH, Zhong DB, Wang YP, Xu XQ, Zhu LH, Tabien RE, Stansel JW, Ying CS. A “defeated” rice resistance gene acts as a QTL against a virulent strain of Xanthomonas oryzae pv. oryzae. Mol Gen Genet, 1999, 261: 58-63[本文引用:1]
 , 孙强
, 孙强

{kind=link}