关键词:MAGIC; 水稻白叶枯病; QTL; 全基因组关联分析; 水稻 Mapping of QTLs for Bacterial Blight Resistance and Screening of Resistant Materials Using MAGIC Populations of Rice CHEN Tian-Xiao1,2, ZHU Ya-Jun3, MI Xue-Fei3, CHEN Kai3, MENG Li-Jun3, ZUO Shi-Min1,*, XU Jian-Long2,3,4,* 1Key Laboratory of Plant Functional Genomics of Jiangsu Province / Key Laboratory of Crop Genetics and Physiology of Jiangsu Province, Yangzhou University, Yangzhou 225009, China
2Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
3Agricultural Genomics Institute at Shenzhen, Chinese Academy of Agricultural Sciences, Shenzhen 518210, China
4Shenzhen Institute of Breeding & Innovation, Chinese Academy of Agricultural Sciences, Shenzhen 518120, China
Fund:This study was supported by National High-Tech Research & Development Plan (863 Program) (2014AA10A601), Shenzhen Peacock Plan (20130415095710361), Scientific and Technological Innovation Project of Chinese Academy of Agricultural Sciences, and the Key Research Plan of Modern Agriculture of Jiangsu Province (BE2015363) AbstractThree genetically interconnected multi-parents advanced generation inter-cross (MAGIC) population, including two populations (DC1 and DC2) derived from four parents and one population from eight parents (DC3) were used to detect QTLs for resistance to two strains, a weak virulent C2 and a strong virulent GD-V of Xanthomonas oryzaepv. oryzae( Xoo) and to screen resistant breeding materials. Most parents were resistant to C2 and susceptible to GD-V. Transgressive segregations of lesion length for the two strains were observed in the three MAGIC populations and showed continuous distributions. A total of seven QTLs affecting lesion length of two strains were detected. Most QTLs showed quantitative resistance and obvious genetic background effect. Among the seven QTLs, QBbr11-1and QBbr11-2 had less genetic background effect, which is valuable in rice breeding for disease resistance. Eight resistant lines pyramiding different QTLs were screened from the three MAGIC populations, indicating the combination of qualitative resistance gene and quantitative resistance gene can significantly improve resistance level. The eight resistant breeding lines could be used as resistant donors in rice breeding for resistance. The results indicated that the MAGIC populations are ideal material for genetic study and marker-assisted breeding, showing a tight integration of genetic research and breeding application in rice.
Keyword:Multi-parent Advanced Generation Inter-Crosses (MAGIC); Rice bacterial blight; Quantitative trait loci (QTL); Genome-wide association study (GWAS); Rice Show Figures Show Figures
表2 亲本对不同菌系的抗性差异比较 Table 2 Comparison of resistance to the Xoo strains among eight parents
亲本1) Parent 1)
C2
GD-V
病斑长度 Lesion length (cm)
显著水平2)Significance level 2)
病斑长度 Lesion length (cm)
显著水平2)Significance level 2)
5%
1%
5%
1%
A
2.6
c
CD
20.7
a
A
B
6.8
b
B
16.0
b
B
C
1.9
cd
CD
13.0
c
C
D
1.3
d
D
9.6
d
D
E
13.5
a
A
16.5
b
B
F
1.0
d
D
12.8
c
C
G
1.4
d
D
9.5
d
D
H
1.7
d
CD
10.7
cd
CD
1)Codes of A-H represent varieties given in Table 1.2) Different capital or small letter are significantly different at the 0.01 or 0.05 probability levels, respectively. 1) 亲本A~H的代号见表1。2) 大写或者是小写字母不同表示其差异在0.01或0.05水平上显著。
表2 亲本对不同菌系的抗性差异比较 Table 2 Comparison of resistance to the Xoo strains among eight parents
表3 Table 3 表3(Table 3)
表3 MAGIC群体接种白叶枯病菌后的病斑长度 Table 3 Lesion length (cm) of the MAGIC populations after inoculating two Xoo races
菌系 Race
DC1
DC2
DC3
均值± 标准差 Mean± SD
变异范围 Range
均值± 标准差 Mean± SD
变异范围 Range
均值± 标准差 Mean± SD
变异范围 Range
C2
2.3± 2.3
0.5-15.5
4.4± 4.2
0.6-21.5
4.2± 4.7
0.4-21.7
GD-V
11.6± 3.8
3.8-23.8
12.0± 3.7
3.6-22.7
11.8± 3.8
1.3-24.0
表3 MAGIC群体接种白叶枯病菌后的病斑长度 Table 3 Lesion length (cm) of the MAGIC populations after inoculating two Xoo races
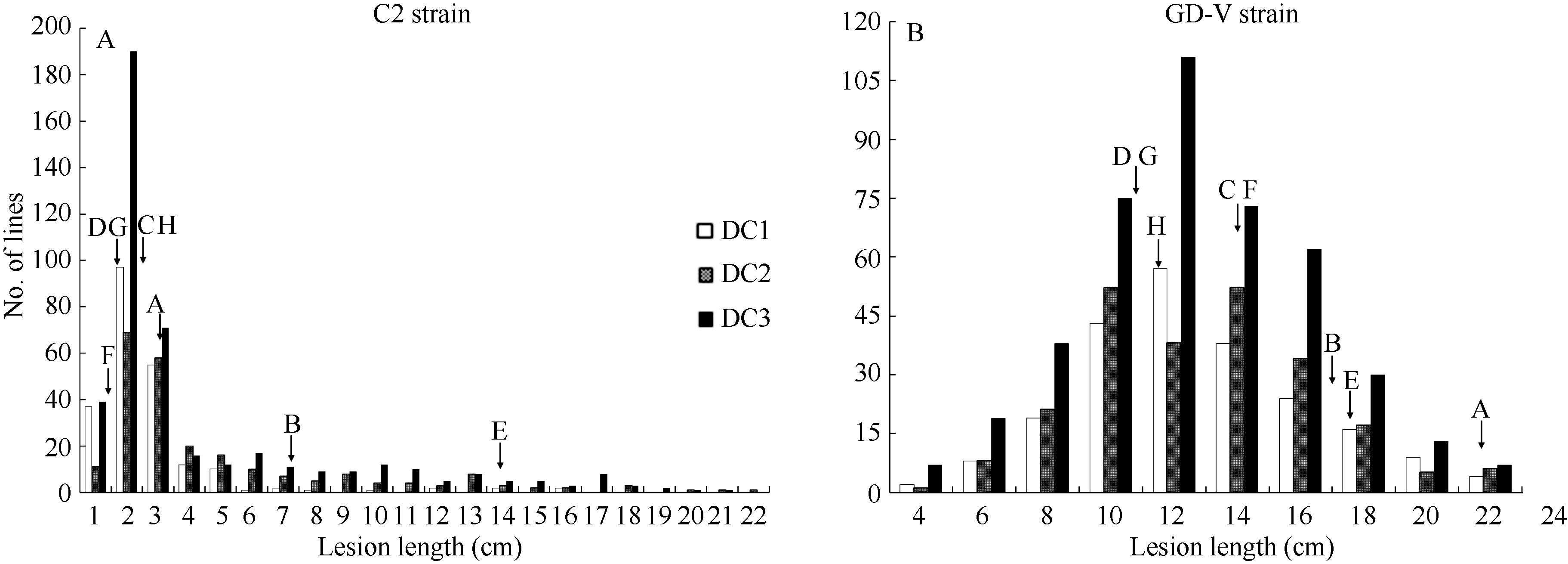
图1 MAGIC群体接种C2 (A)和GD-V (B)菌株的病斑长度频率分布亲本A~H的代号见表1。Codes of A-H represent varieties given in Table 1.Fig. 1 Frequency distributions of lesion length caused by Xoo strain, C2 (A) and GD-V (B) in the MAGIC populations
表4 Table 4 表4(Table 4)
表4 MAGIC群体接种不同菌系病斑长度的方差组成 Table 4 Variance component estimates for multiple factors of lesion length caused by Xoo in the MAGIC populations
群体 Population
变异来源 Source
自由度 df
平方和 SS
均方 MS
F值 F-value
P值 P-value
解释度 R2(%)
DC1
重复Replication
1
3.7
3.7
1.4
0.24
0.01
菌株Race (R)
1
19020.2
19020.2
7184.8
< .0001
66.40
基因型Genotype (G)
220
5030.7
22.9
8.6
< .0001
17.56
基因型× 菌株G× R
220
3424.4
15.6
5.9
< .0001
11.95
误差Error
441
1167.5
2.6
4.08
总计Total
883
28646.5
DC2
重复Replication
1
10.4
10.4
3.2
0.08
0.03
菌株Race (R)
1
13969.4
13969.4
4244.4
< .0001
45.78
基因型Genotype (G)
240
11601.4
48.3
14.7
< .0001
38.02
基因型× 菌株G× R
240
3352.0
14.0
4.2
< .0001
10.98
误差Error
481
1583.1
3.3
5.19
总计Total
963
30516.2
DC3
重复Replication
1
6.2
6.2
3.0
0.09
0.01
菌株Race (R)
1
25803.2
25803.2
12367.3
< .0001
43.11
基因型Genotype (G)
454
23489.1
51.7
24.8
< .0001
39.24
基因型× 菌株G× R
454
8659.9
19.1
9.1
< .0001
14.47
误差Error
909
1896.5
2.1
3.17
总计Total
1819
59854.9
表4 MAGIC群体接种不同菌系病斑长度的方差组成 Table 4 Variance component estimates for multiple factors of lesion length caused by Xoo in the MAGIC populations
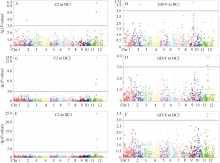
图3 全基因组关联分析定位影响MAGIC群体对2个白叶枯病菌的抗性QTLA、C、E表示混合线性模型下DC1、DC2和DC3群体中与C2菌系抗性显著关联标记的曼哈顿散点图; B、D、F表示混合线性模型下DC1、DC2和DC3群体中与GD-V菌系抗性显著关联标记的曼哈顿散点图。蓝色实线表示全基因组关联分析中的显著性阈值。Fig. 3 Genome-wide association analysis of QTLs underlying resistance to two bacterial blight strains in MAGIC populationsA, C, E: Manhattan plots (MLM) showing genome-wide association study (GWAS) with bacterial blight resistance to strain C2 in DC1, DC2, and DC3 MAGIC populations, respectively. B, D, F: Manhattan plots (MLM) showing GWAS with bacterial blight resistance to strain GD-V in DC1, DC2, and DC3 MAGIC populations, respectively. Blue solid line indicates the genome-wide significance threshold.
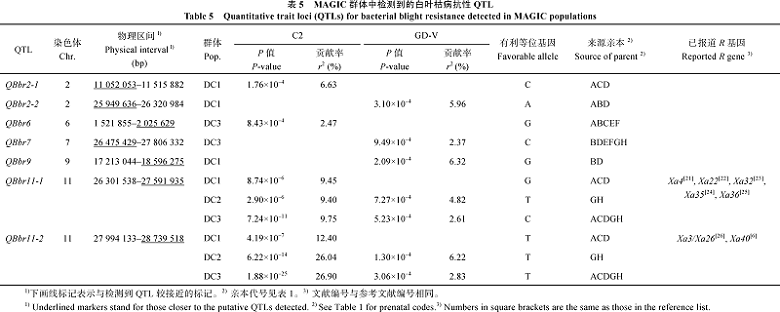
表5 MAGIC 群体中检测到的白叶枯病抗性QTL Table 5 Quantitative tralt locl(QTLs) for bacterial blight resistance detected in MAGIC populations
表6 Table 6 表6(Table 6)
表6 MAGIC群体中筛选出抗病株系的抗性及抗病QTL的分布 Table 6 Lines exhibiting resistance to bacterial blight screened from MAGIC populations and distributions of resistant QTLs
株系 Line
群体 Pop.
病斑长度Lesion length (cm)
携带的QTL QTL carried
QTL数 No. of QTLs
C2
GD-V
L732
DC1
0.60± 0.32 ab
2.30± 0.78 ab
QBbr2-2, QBbr9, QBbr11-2
3
L1273
DC2
0.86± 0.37 ab
3.11± 1.19 bc
QBbr11-1, QBbr11-2
2
L1897
DC3
0.60± 0.26 ab
1.28± 0.89 a
QBbr6, QBbr7, QBbr11-1, QBbr11-2
4
L1602
DC3
1.77± 1.04 c
2.90± 0.41 bc
QBbr11-1, QBbr11-2
2
L2058
DC3
0.53± 0.23 a
3.63± 1.60 c
QBbr6, QBbr7, QBbr11-1
3
L1702
DC3
0.68± 0.28 ab
3.64± 1.19 c
QBbr6, QBbr7, QBbr11-1
3
L1959
DC3
1.08± 0.43 b
3.75± 2.10 c
QBbr6, QBbr7, QBbr11-1
3
L1674
DC3
0.39± 0.26 a
3.82± 1.18 c
QBbr6, QBbr7, QBbr11-1
3
Values followed by different letters are significantly different at the 0.05 probability level. 标以不同字母的值在P=0.05水平上差异显著。
表6 MAGIC群体中筛选出抗病株系的抗性及抗病QTL的分布 Table 6 Lines exhibiting resistance to bacterial blight screened from MAGIC populations and distributions of resistant QTLs
4 结论在遗传上相互关联的2个四亲本群体和1个八亲本群体中观察到对白叶枯病弱毒菌系C2和强毒菌系GD-V的抗性超亲分离。共检测到影响水稻白叶枯病抗性的7个QTL, 大多QTL均表现数量抗性, 抗性表达存在明显的遗传背景效应。QBbr11-2对C2表现质量抗性, 对GD-V表现一定的抗性残余效应。QBbr11-1和QBbr11-2受遗传背景影响较小, 具有一定的育种应用价值。从3个群体筛选出8份不同抗病QTL聚合的抗病材料, 可用作抗病育种的中间抗源。本研究证实了水稻多亲本群体既是遗传研究群体也是理想的育种群体, 可以实现基因定位与育种应用的有机结合。 致谢: 感谢中国农业科学院作物科学研究所水稻分子遗传与分子育种实验室周永力老师和王明明同学帮助培养菌株, 张强、张建、王小倩等同学帮助田间接种和病斑调查。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Mew TW. Current status and future prospects of research on bacterial blight of rice. Annu Rev Phytopathol, 1987, 25: 359-382[本文引用:1]
[2]
ChenS, LiuX, ZengL, Ou-Yang D M, Yang J, Zhu X. Genetic analysis and molecular mapping of a novel recessive gene xa34(t) for resistance against Xanthomonas oryzae pv. oryzae. Theor Appl Genet, 2011, 122: 1331-1338[本文引用:1]
[3]
Niño-Liu DO, Ronald PC, Bogdanove AJ. Xanthomonas oryzae pathovars: model pathogens of a model crop. Mol Plant Pathol, 2006, 7: 303-324[本文引用:1]
[4]
虞玲锦, 张国良, 丁秀文, 高雨, 谢寅峰. 水稻抗白叶枯病基因及其应用研究进展. 植物生理学报, 2012, 48: 223-231Yu LJ, Zhang GL, Ding XW, GaoY, Xie YF. Progress in identification and application of resistance genes to bacterial blight. Plant Physiol J, 2012, 48: 223-231 (in Chinese with English abstract)[本文引用:1]
[5]
罗利军, 梅捍卫, 赵新华, 钟代彬, 王一平, 余新桥, 应存山. 水稻白叶枯病抗性基因定位及其小种专化性. 中国科学, 1998, 28: 536-541Luo LJ, Mei HW, Zhao XH, Zhong DB, Wang YP, Yu XQ, Ying CS. Mapping of resistance genes to bacterial blight of rice and their race specificity in rice. Chin Sci, 1998, 28: 536-541 (in Chinese)[本文引用:1]
[6]
Kim SM, Suh JP, QinY, Noh TH, Reinke RF, Jena KK. Identification and fine-mapping of a new resistance gene, Xa40, conferring resistance to bacterial blight races in rice (Oryza sativa L. ). Theor Appl Genet, 2015, 128: 1933-1943[本文引用:2]
[7]
章琦. 中国杂交水稻白叶枯病抗性的遗传改良. 中国水稻科学, 2009, 23: 111-119ZhangQ. Genetics and improvement of resistance to bacterial blight in hybrid rice in China. Chin J Rice Sci, 2009, 23: 111-119 (in Chinese with English abstract)[本文引用:2]
[8]
Khan MA, NaeemM, IqbalM. Breeding approaches for bacterial leaf blight resistance in rice (Oryza sativa L. ), current status and future directions. Eur J Plant Pathol, 2014, 139: 27-37[本文引用:1]
Ea V DB, Jones JD. Plant disease-resistance proteins and the gene-for-gene concept. Trends Biolchem Sci, 1998, 23: 454-456[本文引用:1]
[11]
Li ZK, Luo LJ, Mei HW, Paterson AH, Zhao XH, Zhong DB, Wang YP, Yu XQ, ZhuL, TabienR, Stansel JW, Ying CS. A “defeated” rice resistance gene acts as a QTL against a virulent strain of Xanthomonas oryzae pv. oryzae. Mol Gen Genet, 1999, 261: 58-63[本文引用:1]
[12]
Zhao KY, Tung CW, Eizenga GC, Wright MH, Ali ML, Price AH, Norton GJ, Islam MR, ReynoldsA, MezeyJ, McClung A M, Bustamante C D, McCouch S R. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat Commun, 2011, 2: 1-10[本文引用:1]
[13]
Meng LJ, Zhao XQ, PonceK, LeungH, Ye GY. QTL mapping for agronomic traits using multi-parent advanced generation inter-cross (MAGIC) populations derived from diverse elite indica rice lines. Field Crops Res, 2016, 189: 19-42[本文引用:2]
[14]
Kauffman HE, Reddy A P K, Hsieh S P Y, Merca S D. Improved technique for evaluating resistance of rice varieties to Xanthomonas oryzae. Plant Dis Rep, 1973, 57: 537-541[本文引用:1]
[15]
BradburyP, ZhangZ, KroonD, Casstevens TY, BucklerE. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1]
[16]
ValdarW, FlintJ, MottR. Simulating the collaborative cross: Power of quantitative trait loci detection and mapping resolution in large sets of recombinant inbred strains of mice. Genetics, 2006, 172: 1783-1797[本文引用:1]
[17]
CavanaghC, MorellM, MackayI, PowellW. From mutations to MAGIC, resources for gene discovery, validation and delivery in crop plants. Curr Opin Plant Biol, 2008, 11: 215-221[本文引用:1]
[18]
Huang BE, Verbyla KL, Verbyla AP, RaghavanC, Singh VK, GaurP, LeungH, Varshney RK, Cavanagh CR. MAGIC populations in crops: current status and future prospects. Theor Appl Genet, 2015, 128: 999-1017[本文引用:1]
[19]
Band illoN, RaghavanC, Muyco PA, Sevilla M A L, Lobina I T, Dilla-Ermita C J, Tung C W, McCouch S, Thomson M, Mauleon R, Singh R K, Gregorio G, Redoña E, Leung H. Multi-parent advanced generation inter-cross (MAGIC) populations in rice: progress and potential for genetics research and breeding. Rice, 2013, 6: 1-15[本文引用:1]
[20]
LeungH, RaghavanC, ZhouB, OlivaR, Choi IR, LacorteV, Jubay ML, Cruz CV, GregorioG, Singh RK, Ulat VJ, Borja FN, MauleonR, Alexand rov NN, McNally K L, Hamilton R S. Allele mining and enhanced genetic recombination for rice breeding. Rice, 2015, 8: 1-11[本文引用:1]
[21]
SunX, YangZ, WangS, ZhangQ. Identification of a 47-kb DNA fragment containing Xa4, a locus for bacterial blight resistance in rice. Theor Appl Genet, 2003, 106: 683-687[本文引用:1]
[22]
Wang CT, Tan MP, XuX, Wen GS, Zhang DP, Lin XH. Localizing the bacterial blight resistance gene, Xa22(t), to a 100-kilobase bacterial artificial chromosome. Phytopathology, 2003, 93: 1258-1262[本文引用:1]
[23]
郑崇珂, 王春连, 于元杰, 梁云涛, 赵开军. 水稻抗白叶枯病新基因Xa32(t)的鉴定和初步定位. 作物学报, 2009, 35: 1173-1180Zheng CK, Wang CL, Yu YJ, Liang YT, Zhao KJ. Identification and molecular mapping of Xa32(t), a novel resistance gene for bacterial blight (Xanthomonas oryzae pv. oryzae) in rice. Acta Agron Sin, 2009, 35: 1173-1180 (in Chinese with English abstract)[本文引用:1]
[24]
郭嗣斌, 张端品, 林兴华. 小粒野生稻抗白叶枯病新基因的鉴定与初步定位. 中国农业科学, 2010, 43: 2611-2618Guo SB, Zhang DP, Lin XH. Identification and mapping of a novel bacterial blight resistance gene Xa35(t) originated from oryza minuta. Sci Agric Sin, 2010, 43: 2611-2618 (in Chinese with English abstract)[本文引用:1]
[25]
苗丽丽, 王春连, 郑崇珂, 车晋英, 高英, 温义昌, 李贵全, 赵开军. 水稻抗白叶枯病新基因的初步定位. 中国农业科学, 2010, 43: 3051-3058Miao LL, Wang CL, Zheng CK, Che JY, GaoY, Wen YC, Li GQ, Zhao KJ. Molecular mapping of a new gene for resistance to rice bacterial blight. Sci Agric Sin, 2010, 43: 3051-3058 (in Chinese with English abstract)[本文引用:1]
[26]
XiangY, Cao YL, Xu CG, Li XH, Wang SP. Xa3, conferring resistance for rice bacterial blight and encoding a receptor kinase-like protein, is the same as Xa26. Theor Appl Genet, 2006, 113: 1347-1355[本文引用:1]
[27]
WangY, ZhangQ, Zheng TQ, Cui YR, Zhang WZ, Xu JL, Li ZK. Drought-tolerance QTLs commonly detected in two sets of reciprocal introgression lines in rice. Crop & Pasture Sci, 2014, 65: 171-184[本文引用:1]
[28]
WangY, Zang JP, SunY, AliJ, Xu JL, Li ZK. Background-independent quantitative trait loci for drought tolerance identified using advanced backcross introgression lines in rice. Crop Sci, 2013, 53: 430-441[本文引用:1]
[29]
Cheng LR, WangY, Meng LJ, HuX, Cui YR, SunY, Zhu LH, AliJ, Xu JL, Li ZK. Identification of salt-tolerant QTLs with strong genetic background effect using two sets of reciprocal introgression lines in rice. Genome, 2012, 55: 45-55[本文引用:1]
[30]
杨静, 孙勇, 程立锐, 周政, 王韵, 朱苓华, 苍晶, 徐建龙, 黎志康. 利用双向导入系群体检测遗传背景对耐盐QTL定位的影响. 作物学报, 2009, 35: 974-982YangJ, SunY, Cheng LR, ZhouZ, WangY, Zhu LH, CangJ, Xu JL, Li ZK. Genetic background effect on QTL mapping for salt tolerance revealed by a set of reciprocal introgression line populations in rice. Acta Agron Sin, 2009, 35: 974-982 (in Chinese with English abstract)[本文引用:1]
[31]
BanerjeeD, ZhangX, Bent AF. The leucine-rich repeat domain can determine effective interaction between RPS2 and other host factors in Arabidopsis RPS2-mediated disease resistance. Genetics, 2001, 158: 439-450[本文引用:1]
[32]
徐建龙, 林贻滋, 张炳林, 翁锦屏. 水稻白叶枯病抗性基因Xa-21的初步利用. 浙江农业学报, 1996, 8: 70-73Xu JL, Lin YZ, Zhang BL, Weng JP. Study on utilization of Xa-21 gene resistant to bacterial blight in rice. Acta Agric Zhejianggensis, 1996, 8: 70-73 (in Chinese with English abstract)[本文引用:1]
[33]
Sun XL, Gao YL, Yang ZF, Xu CG, Li XH, Wang SP, Zhang QF. Xa26, a gene conferring resistance to Xanthomonas oryzae pv. oryzae in rice, encodes an LRR receptor kinase-like protein. Plant J, 2004, 37: 517-527[本文引用:1]
[34]
于彦春, 滕胜, 曾大力, 董国军, 钱前, 黄大年, 朱立煌. 水稻抗白叶枯病微效QTL的定位分析. 中国水稻科学, 2003, 17: 315-318Yu YC, TengS, Zeng DL, Dong GJ, QianQ, Huang DN, Zhu LH. Analysis of QTLs for resistance to rice bacterial bight. Chin J Rice Sci, 2003, 17: 315-318 (in Chinese with English abstract)[本文引用:1]
[35]
Wang CM, Su CC, Zhai HQ, Wan JM. Identification of QTLs underlying resistance to a virulent strain of Xanthomonas oryzae pv. oryzae in rice cultivar DV85. Field Crops Res, 2005, 91: 337-343[本文引用:1]
[36]
Li ZK, ArifM, Zhong DB, Fu BY, Domingo-ReyJ, AliJ, Vijayakumar C H M, Yu S B, Khush G S. Complex genetic networks underlying the defensive system of rice (Oryza sativa L. ) to Xanthomonas oryzae pv. oryzae. Proc Natl Acad Sci USA, 2006, 103: 7994-7999[本文引用:1]
[37]
ZhangF, Xie XW, Xu MR, Wang WS, Xu JL, Zhou YL, Li ZK. Detecting major QTL associated with resistance to bacterial blight using a set of rice reciprocal introgression lines with high density SNP markers. Plant Breed, 2015, 134: 286-292[本文引用:2]
[38]
Zhou YL, Uzokwe V N E, Zhang C H, Cheng L R, Wang L, Chen K, Gao X Q, Sun Y, Chen J J, Zhu L H, Zhang Q, Ali J, Xu J L, Li Z K. Improvement of bacterial blight resistance of hybrid rice in China using the Xa23 gene derived from wild rice (Oryza rufipogon). Crop Protection, 2011, 30: 637-644[本文引用:1]
[39]
Lee SW, Choi SH, Han SS, Lee DG, Lee BY. Distribution of Xanthomonas oryzae pv. oryzae strains virulent to Xa21 in Korea. Phytopathology, 1999, 89: 928-933[本文引用:1]
[40]
Shanti ML, George M L C, Cruz C M V, Bernardo M A, Nelson R J, Leung H, Reddy J N, Sridhar R. Identification of resistance genes effective against rice bacterial blight pathogen in eastern India. Plant Dis, 2001, 85: 506-512[本文引用:1]
[41]
曾列先, 黄少华, 伍尚忠. IRBB21 (Xa21)对广东稻白叶枯病菌5个小种的抗性反应. 植物保护学报, 2002, 29: 97-100Zeng LX, Huang SH, Wu SZ. Resistance of IRBB21 (Xa21) to five races of bacterial blight in Guangdong. Acta Phytophyl Sin, 2002, 29: 97-100 (in Chinese)[本文引用:1]
[42]
Gu KY, YangB, Tian DS, Wu LF, Wang DJ, SreekalaC, YangF, Chu ZQ, Wang GL, White FF, Yin ZC. R gene expression induced by a type-III effector triggers disease resistance in rice. Nature, 2005, 435: 1122-1125[本文引用:1]
 , 徐建龙
, 徐建龙


{kind=link}
{kind=link}
{kind=link}