关键词:海岛棉; 抗黄萎病; 未知基因( GbVWR); 克隆; 基因表达 Cloning and Expression Analysis of a Functional Gene GbVWR Induced by Verticillium dahliae in Gossypium barbadense ZHANG Li-Jia**, ZHANG Yan**, RONG Wei, YANG Jun, ZHANG Gui-Yin, WU Li-Qiang, LI Zhi-Kun, WU Jin-Hua, MA Zhi-Ying, WANG Xing-Fen* North China Key Laboratory for Crop Germplasm Resources of Education Ministry / Key Laboratory for Crop Germplasm Resources of Hebei / Agricultural University of Hebei, Baoding 071001, China Fund:This study was supported by the Natural Science Foundation of Hebei Province (C2013204141), the Special Research Found for the Doctoral Program of Higher Education (20131302120002), and the Science and Technology Support Project of Hebei Province (14226308D). Abstract Verticillium dahliae is a destructive, soil-borne fungal pathogen that causes severe losses in cotton yield and fiber quality. Mining functional genes related to resistance against V. dahliae will benefit efforts to genetically improve crop plants. In this study, we identified a gene that involved in cotton defense against V. dahliaebased on screening the full-length cDNA library and suppression subtractive hybridization library (SSH) induced by V. dahliae in Gossypium barbadenseand Gossypium. hirsutum, respectively. Sequence analysis indicated that there was no any annotation in NCBI database, and we named the sequence from G. barbadense as GbVWR. We characterized GbVWR gene and analyzed its expression. The full length cDNA of GbVWR was 520 bp including a 198 bp open reading frame (ORF), encoding 65 amino acid residues. Bioinformatic analyses suggested that GbVWR belonged to secretory protein and tis theoretical isoelectric point was 5.32. Using pET-32a(+) as a fused expression vector, a recombinant plasmid pET32a-GbVWR was constructed. The recombinant protein was induced in Escherichia coliBL21 (DE3) with 1.0 mmol L-1 IPTG then GbVWR could express about 7 kD protein in E. coliBL21 (DE3). In addition, diverse cis-acting promoter elements involved in fungal elicitor response, hormone response, wound-response, and flavonoid biosynthetic gene regulation were discovered in the promoter region of GbVWR. qPCR analysis showed that expression level of GbVWR was the highest in roots, and was significantly induced by V. dahliae. Besides, GbVWR could also be induced by SA, ET, and GA treatments, respectively. In conclusion, GbVWR is a new functional gene, which involved in multiple signal pathways in cotton defense response to Verticillium wilt.
Keyword: Gossypium barbadense; Verticillium wilt resistance; GbVWR; Clone; Gene expression Show Figures Show Figures
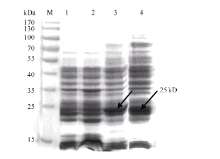
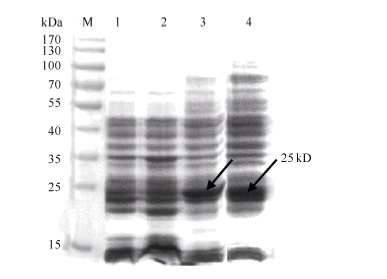
图2 重组蛋白pET32a-GbVWR表达产物SDS-PAGE分析箭头所指为目的蛋白条带; M: 蛋白分子量标准; 1: pET32a(+)空载体37℃下诱导9 h; 2: BL21(DE3)空菌株37℃下诱导9 h; 3~4: pET32a-GbVWR 37℃下分别诱导9 h和24 h。Fig. 2 SDS-PAGE analysis of the expressed product of recombinant pET32a-GbVWRArrows indicate the target protein bands; M: protein molecular weight standard; 1: strains harbouring pET-32a(+) after induction for 9 hours at 37℃; 2: strains harbouring BL21 (DE3) after induction for 9 hours at 37℃; 3-4: total proteins of engineering bacteria strains after induction for 9 and 24 hours at 37℃, respectively.
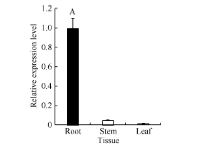
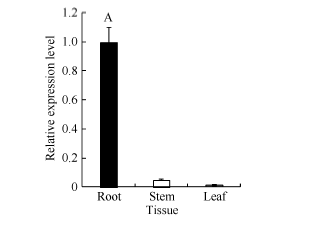
图3GbVWR在海岛棉品系Pima 90-53不同组织中的RT-PCR分析同一处理时间内, “ A” 表示在0.01水平上存在极显著差异。Fig. 3 Tissue-specific expression analysis pattern of GbVWR by qRT-PCR in the G. barbadensePima 90-53“ A” means the significant difference at the 0.01 probability level at the same treatment.
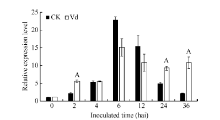
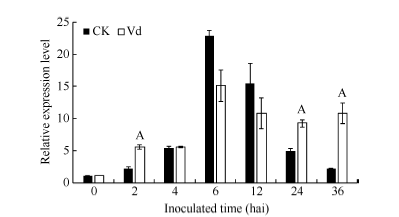
图4GbVWR在棉花黄萎病菌胁迫下的RT-PCR分析同一处理时间内, “ A” 表示在0.01水平上存在极显著差异。Fig. 4 Expression analysis of GbVWRunder Verticillium dahliaestress“ A” means the significant difference at the 0.01 probability level at the same treatment.
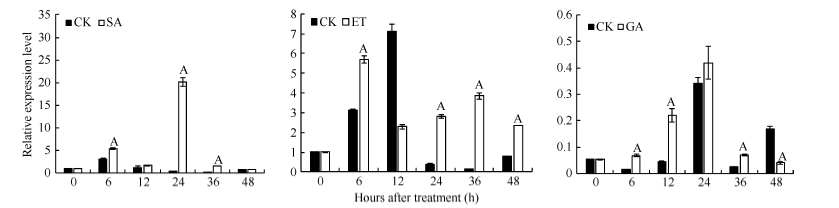
图5 外源激素处理下GbVWR的表达模式分析同一处理时间内, “ A” 表示在0.01水平上存在极显著差异, “ a” 表示在0.05水平上存在显著差异。Fig. 5 Expression analysis of VWRafter treatment by hormones“ A” means the significant difference at the 0.01 probability level at the same treatment, “ a” means the significant difference at the 0.05 probability level.
Zhang JF, SanogoS, FlynnR, Baral JB, BajajS, Hughs SE, Percy RG. Germplasm evaluation and transfer of Verticillium wilt resistance from Pima ( Gossypium barbadense) to upland cotton( G. hirsutum). Euphytica, 2012, 187: 147-160[本文引用:2]
[4]
徐理, 朱龙付, 张献龙. 棉花抗黄萎病机制研究进展. 作物学报, 2012, 38: 1553-1560XuL, Zhu LF, Zhang XL. Research on resistance mechanism of cotton to Verticillium wilt. Acta Agron Sin, 2012, 38: 1553-1560 (in Chinese with English abstract)[本文引用:2]
[5]
SanogoS, Zhang JF. Resistance sources, resistance screening techniques and disease management for Fusarium wilt in cotton. Euphytica, 2016, 207: 255-271[本文引用:1]
[6]
WilhelmS, Sagen JE, TietzH. Resistance to Verticillium wilt in cotton: sources, techniques of identification, inheritance trends, and the resistance potential of multilinecultivars. Phytopathology, 1974, 64: 924-931[本文引用:1]
[7]
Bell AA. Verticillium wilt. In: Hillocks R J ed. Cotton Diseases. Wallingford, UK: CAB Intern, 1992. pp 87-126[本文引用:2]
[8]
BellAA. Verticillium wilt. In: Kirkpatrick T L, Rothrock C S, eds. Compendium of Cotton Diseases. Amer. St. Paul, MN: Phytopathol. Soc, 2001. pp 28-31[本文引用:1]
[9]
Cai YF, He XH, Mo JC, SunQ, Yang JP, Liu JG. Molecular research and genetic engineering of resistance to Verticillium wilt in cotton. Afr J Biotechnol, 2009, 8: 7363-7372[本文引用:1]
[10]
Wendel JF, Cronn RC. Polyploidy and the evolutionary history of cotton. Adv Agron, 2003, 78: 139-186[本文引用:1]
[11]
Zhang JF, SanogoS, FlynnR, Baral JB, BajajS, Hughs SE. Germplasm evaluation and transfer of Verticillium wilt resistance from Pima ( Gossypium barbadense) to Upland cotton( G. hirsutum). Euphytica, 2012, 187: 147-160[本文引用:2]
[12]
Zhou HP, FangH, SanogoS, Hughs SE, Jones DC, Zhang JF. Evaluation of Verticillium wilt resistance in commercial cultivars and advanced breeding lines of cotton. Euphytica, 2014, 196: 437-448[本文引用:2]
[13]
Zhang JF, Percy RG, McCarty J J. Introgression genetics and breeding between Upland and Pima cotton: a review. Euphytica, 2014, 198: 1-12[本文引用:1]
[14]
ZhangY, Wang XF, YangS, Chi JN, Zhang GY, Ma ZY. Cloning and characterization of a Verticillium wilt resistance gene from Gossypium barbadense and functional analysis in Arabidopsis thaliana. Plant Cell Rep, 2011, 30: 2085-2096[本文引用:1]
[15]
Zhang BL, Yang YW, Chen TZ, Yu WG, Liu TL, Li HJ, Fan XH, Ren YZ, Shen DY, LiuL, Duo DL, Chang YH. Island cotton Gbve1 gene encoding a receptor-like protein confers resistance to both defoliating and non-defoliating isolates of Verticillium dahliae. PLoS One, 2012, 7(12): e51091[本文引用:1]
[16]
Gao XQ, WheelerT, Li ZH, Kenerley CM, HeP, Shan LB. Silencing GhNDR1and GhMKK2compromises cotton resistance to Verticillium wilt. Plant J, 2011, 66: 293-305[本文引用:1]
[17]
ZhangY, Wang XF, Li YY, Wu LZ, Zhou HM, Zhang GY, Ma ZY. Ectopic expression of a novel Ser/Thr protein kinase from cotton ( Gossypium barbadense), enhances resistance to Verticillium dahliaeinfection and oxidative stress in Arabidopsis. Plant Cell Rep, 2013, 32: 1703-1713[本文引用:1]
[18]
Mo HJ, Wang XF, ZhangY, Zhang GY, Zhang JF, Ma ZY. Cotton polyamine oxidase is required for spermine and camalexin signalling in the defence response to Verticillium dahliae. Plant J, 2015, 83: 962-975[本文引用:2]
[19]
杨君, 张艳, 王伟巧, 吴金华, 王国宁, 马峙英, 王省芬. 海岛棉 GbHyPRP1克隆及其转基因拟南芥抗黄萎病验证. 植物资源学报, 2015, 16: 594-602YangJ, ZhangY, Wang WQ, Wu JH, Wang GN, Ma ZY, Wang XF. Cloning of GbHyPRP1 from Gossypium barbadense and validation of Verticillium wilt resistance in transgenic Arabidopsis. J Plant Genet Resour, 2015, 16: 594-602 (in Chinese with English abstract)[本文引用:1]
[20]
XuL, Zhu LF, Tu LL, Yuan DJ, JinL, LongL, Zhang XL. Lignin metabolism has a central role in the resistance of cotton to the wilt fungus Verticillium dahliae as revealed by RNA-Seq- dependent transcriptional analysis and histochemistry. J Exp Bot, 2011, 62: 5607-5621[本文引用:1]
[21]
ZhangY, Wang XF, Ding ZG, MaQ, Zhang GR, Zhang SL, Li ZK, Wu LQ, Zhang GY, Ma ZY. Transcriptome profiling of Gossypium barbadense inoculated with Verticillium dahliae provides a resource for cotton improvement. BMC Genom, 2013, 14: 637[本文引用:3]
LiuX, ZhaoB, Zheng HJ, HuY, LuG, Yang CQ, Chen JD, Chen JJ, Chen DY, ZhangL, ZhouY, Wang LJ, Guo WZ, Bai YL, Ruan JX, Shang-Guan X X, Mao Y B, Shan C M, Jiang J P, Zhu Y Q, Jin L, Kang H, Chen S T, He X L, Wang R, Wang Y Z, Chen J, Wang L J, Yu S T, Wang B Y, Wei J, Song S C, Lu X Y, Gao Z C, Gu W Y, Deng X, Ma D, Wang S, Liang W H, Fang L, Cai C P, Zhu X F, Zhou B L, Chen Z J, Xu S H, Zhang Y G, Wang S Y, Zhang T Z, Zhao G P, Chen X Y. Gossypium barbadense genome sequence provides insight into the evolution of extra- long staple fiber and specialized metabolites. Sci Rep, 2015, 5: 14139[本文引用:1]
ZhangT, HuY, JiangW, FangL, GuanX, ChenJ, ZhangJ, Saski CA, Scheffler BE, Stelly DM, Hulse-Kemp A M, Wan Q, Liu B, Liu C, Wang S, Pan M, Wang Y, Wang D, Ye W, Chang L, Zhang W, Song Q, Kirkbride R C, Chen X, Dennis E, Llewellyn D J, Peterson D G, Thaxton P, Jones D C, Wang Q, Xu X, Zhang H, Wu H, Zhou L, Mei G, Chen S, Tian Y, Xiang D, Li X, Ding J, Zuo Q, Tao L, Liu Y, Li J, Lin Y, Hui Y, Cao Z, Cai C, Zhu X, Jiang Z, Zhou B, Guo W, Li R, Chen Z J. Sequencing of allotetraploid cotton ( Gossypium hirsutum L. acc. TM-1) provides a resource for fiber improvement. Nat Biotechnol, 2015, 33: 531-537[本文引用:1]
[27]
张纯颖, 王省芬, 张桂寅, 吴立强, 迟吉娜, 李志坤, 马峙英. 黄萎病菌诱导下陆地棉抗病品种SSH文库的EST分析. 棉花学报, 2010, 22: 17-22Zhang CY, Wang XF, Zhang GY, Wu LQ, Chi JN, Li ZK, Ma ZY. EST analysis of suppression subtractive hybridization library from upland cotton resistant cultivar infection by Verticillium dahliae. Cotton Sci, 2010, 22: 17-22 (in Chinese with English abstract)[本文引用:1]
ZhaoP, Zhao YL, JinY, ZhangT, Guo HS. Colonization process of Arabidopsis thalianaroots by a green fluorescent protein-tagged isolate of Verticillium dahliae. Protein Cell, 2014, 5: 94-98[本文引用:1]
[30]
彭姗, 吕学莲, 高峰, 李国英, 李晖. 一种新的棉花黄、枯萎病快速接种方法的研究. 棉花学报, 2008, 20: 174-178PengS, Lyu XL, GaoF, Li GY, LiH. Study on a new rapid inoculation method for Verticillium wilt and Fusarium wilt of cotton. Cotton Sci, 2008, 20: 174-178 (in Chinese with English abstract)[本文引用:1]
[31]
Cheng CX, JiaoC, Singer SD, GaoM, Xu XZ, Zhou YM, LiZ, Fei ZG, Wang YJ, Wang XP. Gibberellin-induced changes in the transcriptome of grapevine ( Vitis labrusca × V. vinifera ) cv. Kyoho flowers. BMC Genom, 2015, 16: 128[本文引用:1]
[32]
陈士林, 王春虎. 钙和赤霉素对棉花种子发芽力及活力的影响. 中国农学通报, 2004, 20(3): 112-113Chen SL, Wang CH. Effects of Ca2+ and GA on the germination and the seed vigour in cotton. Chin Agric Sci Bull, 2004, 20(3): 112-113 (in Chinese with English abstract)[本文引用:1]
[33]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25: 402-408[本文引用:1]
[34]
BariR, Jones J D G. Role of plant hormones in plant defence responses. Plant Mol Biol, 2009 69: 473-488[本文引用:1]
[35]
Clarke JD, Volko SM, LedfordH, Ausubel FM, DongX. Roles of salicylic acid, jasmonic acid, and ethylene in cpr-induced resistance in Arabidopsis. Plant Cell, 2000, 12: 2175-2190[本文引用:1]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}