关键词:烟草; 细胞质雄性不育; ATP合酶; 亚基基因; RNA编辑 Editing Sites in Transcript of Four F0-ATPase Subunit Gene in Tobacco TAO Yao1, WANG Yu1,3, ZHONG Si-Rong1, WU Lin-Min1, XIE Li-Juan1, NIE Ya-Ping1, ZHOU Wei2, WANG Jian-Ge4, LIU Qi-Yuan1,* 1 Key Laboratory of Crop Physiology, Ecology and Genetic Breeding, Ministry of Education / Key Laboratory of Crop Physiology, Ecology and Genetic Breeding of Jiangxi Province / College of Agronomy, Jiangxi Agricultural University, Nanchang 330045, China
2 Hunan Provincial Key Laboratory for Biology and Control of Plant Diseases and Insect Pests, College of Plant Protection, Hunan Agricultural University, Changsha 410128, China
3Agricultural committee of Guizhou province Qianxinanzhou, Xingyi 562400, China
4 College of Landscape Architecture and Art, Jiangxi Agricultural University, Nanchang 330045, China
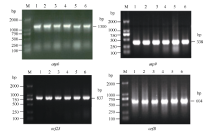
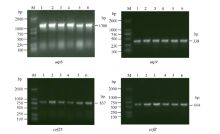
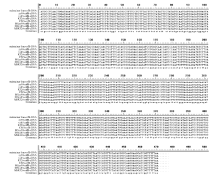
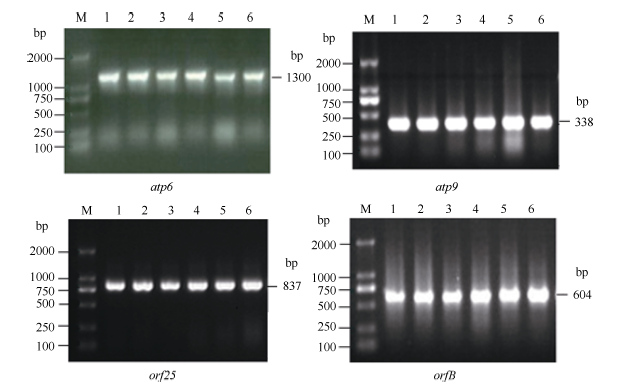
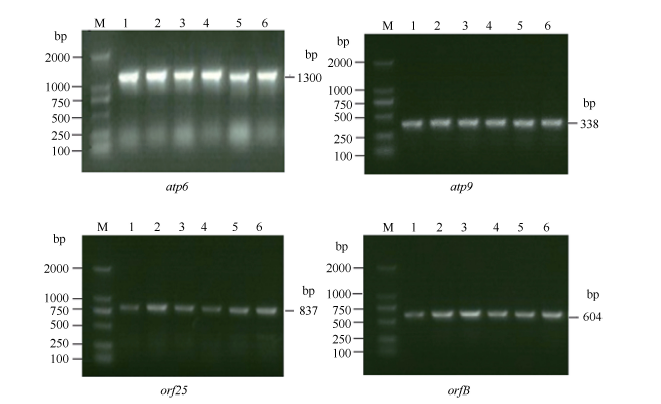
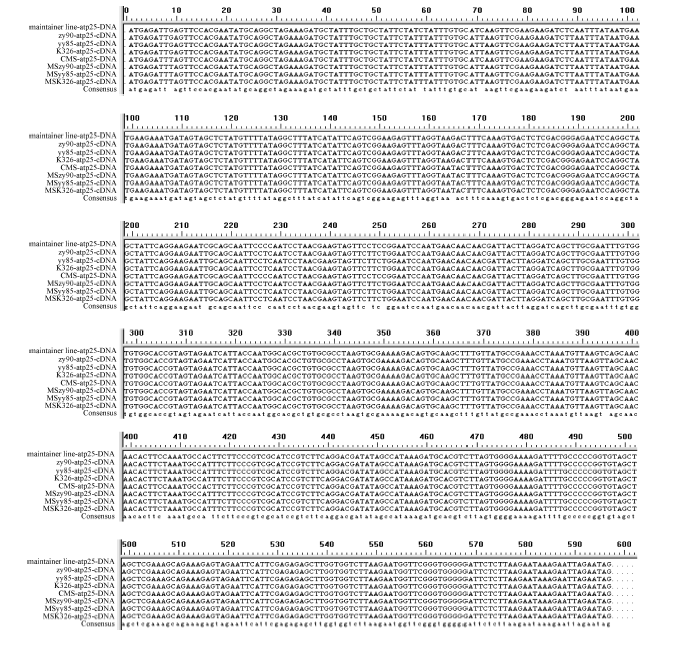
Fund:This study was supported by the National Natural Science Foundation of China (31260350, 31301388), China Postdoctoral Science Foundation (2015T80870, 2014M562109), and Science and Technology Plan Projects of Jiangxi Province Education Department (GJJ13275). AbstractRNA editing exits extensively in mitochondria of higher plants and is one of the most important post-transcriptional regulation methods of gene expression in mitochondrial genomes of higher plants. At the same time, it is an essential process for forming function proteins. RNA editing can induce mutations in mitochondrial genes including nucleotide insertion, substitution, or deletion, which further affects the splicing and processing of primary transcripts, ultimately resulting in cytoplasmic male sterility (CMS). The results of research using multiple species showed that there is an obvious relationship between the four subunit genes of F0-ATPase and CMS. To explore the relationship, we studied RNA editing status of four mitochondrial genes atp6, atp9, orf25, and orfB from three tobacco male sterility lines (MS Zhongyan 90, MS Yunyan 85, MS K326) and their corresponding fertile lines . The four mitochondrial genes atp6, atp9, orf25, and orfB and their cDNA were distinctively amplified by PCR from six tobacco lines. After that, by means of making a comparison between the DNA sequences and the cDNA sequences of target genes to find RNA editing sites. The orf25and orfB gene transcripts had the same RNA editing sites between male sterile and fertile lines. For atp6 gene, RNA editing didn't occur in male sterile lines, while there were six RNA editing sites in fertile lines, which all caused changes in the type of amino acids and there were four editing sites enhancing hydrophobicity of the amino acids. It inferred that the difference of protein's hydrophobicity was most likely to cause CMS. The atp9 gene had ten RNA editing sites in fertile lines, eight of which were the same as those in male sterile lines, while two C→T unique editing sites were absent in male sterile lines, of which one caused changes in amino acid. The nucleotide variations at 223 site of atp9 gene resulted in producing a termination code, which might be the necessary RNA editing to produce normal functional protein. These results suggest that lacking of the unique RNA editing sites might contribute to CMS property in tobacco.
Keyword:Tobacco; Cytoplasmic male sterility; ATPase; Subunit gene; RNA editing Show Figures Show Figures
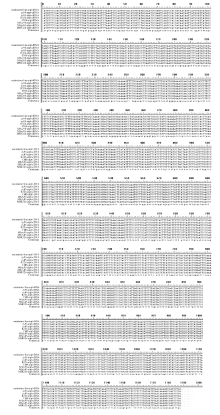
图5 不育系及其保持系烟草中orf25基因的RNA编辑位点及基因突变位点59、71、89、215、227、248、251、395、407、416位点为不育系和保持系orf25基因的RNA编辑位点; 9、165位点为不育系orf25基因的基因突变位点。Fig. 5 RNA editing sites and gene mutation sites oforf25 gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites oforf25 gene between cytoplasmic male sterility and maintainer lines were sites 59, 71, 89, 215, 227, 248, 251, 395, 407, 416; Gene mutation sites oforf25 in cytoplasmic male sterility were sites 9, 165.
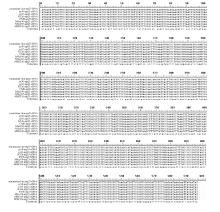
图6 不育系及其保持系烟草中orfB基因的RNA编辑位点及基因突变位点47、58、76、443位点为不育系和保持系orfB基因的RNA编辑位点; 363为不育系orfB基因的基因突变位点。Fig. 6 RNA editing sites and gene mutation sites oforfB gene between cytoplasmic male sterility and maintainer lines in tobaccoRNA editing sites oforfB gene between cytoplasmic male sterility and maintainer lines were sites 47, 58, 76, 443; Gene mutation sites oforfB in cytoplasmic male sterility were site 363.
KellerW, WolfJ, GerberA. Editing of messenger RNA precursors and of tRNAs by adenosine to inosine conversion. FEBS Lett, 1999, 452: 71-76[本文引用:1]
[2]
HieselR, CombettesB, BrennickeA. Evidence for RNA editing in mitochondria of all major groups of land plants except the Bryothyta. Proc Natl Acad Sci USA, 1994, 91: 629-633[本文引用:1]
[3]
刘欣芳, 马骏, 王金艳, 王树坤, 姜敏. 高等植物RNA编辑的研究进展. 辽宁农业科学, 2012, (4): 43-47Liu XF, MaJ, Wang JY, Wang SK, JiangM. Research progress of RNA editing in higher plant. Liaoning Agric Sci, 2012, (4): 43-47 (in Chinese with English abstract)[本文引用:1]
[4]
GiegeP, BrennickeA. RNA editing in Arabidopsis mitochondria effects 441 C to U changes on ORFs. Proc Natl Acad Sci USA, 1999, 96: 15324-15329[本文引用:1]
[5]
LiuZ, Cai XW, Seiler GJ, Jan CC. Interspecific amphiploid-derived alloplasmic male sterility with defective anthers, narrow disc florets and small ray flowers in sunflower. Plant Breed, 2014, 133: 742-747[本文引用:1]
[6]
ChenP, RanS, LiR, Huang ZP, Qian JH, Yu ML, Zhou RY. Transcriptome de novo assembly and differentially expressed genes related to cytoplasmic male sterility in kenaf ( Hibiscus cannabinus L. ). Mol Breed, 2014, 34: 1879-1891[本文引用:1]
[7]
祁建民, 马红勃, 徐建堂, 陈美霞, 周东新, 王涛, 陈顺辉. 烟草细胞质雄性不育系及其保持系的花蕾差异蛋白质分析. 作物学报, 2012, 38: 1232-1239Qi JM, Ma HB, Xu JT, Chen MX, Zhou DX, WangT, Chen SH. Proteomic analysis of bud differentiation between cytoplasmic male-sterile line and maintainer in tobacco. Acta Agron Sin, 2012, 38: 1232-1239 (in Chinese with English abstract)[本文引用:1]
[8]
王震, 范晓静, 张淼, 张芳凝, 李桂东, 马翎健. ATP合成相关基因在小麦BNS不育系育性转换中的差异表达. 作物学报, 2014, 40: 1501-1505WangZ, Fan X J. Zhang M, Zhang FN, Li GD, Ma LJ. Differential expression of ATP synthesis related gene in fertility conversion of wheat BNS male sterile line. Acta Agron Sin, 2014, 40: 1501-1505 (in Chinese with English abstract)[本文引用:1]
[9]
WeiL, Yan ZX, DingY. Mitochondrial RNA editing of F0-ATPase subunit 9 gene ( atp9) transcripts of Yunnan purple rice cytoplasmic male sterile line and its maintainer line. Acta Physiol Plant, 2008, 30: 657-662[本文引用:1]
[10]
Zhao YH, Liao XF, Zhou BJ, Zhao HT, Zhou YY, Zhou RY. Mutation in the coding sequence of atp6are associated with male sterile cytoplasm in kenaf( Hibiscus cannabinus L. ). Euphytica, 2016, 207: 169-175[本文引用:1]
[11]
StahlR, SunS, L'Homme Y, Ketela T, Brown G G. RNA editing of transcripts of a chimeric mitochondrial gene associated with cytoplasmic male-sterility in Brassica. Nucl Acids Res, 1994, 22: 2109-2113[本文引用:1]
魏磊, 费震江, 丁毅. 紫稻( Oryza sativaL . )线粒体ATP合成酶 atp6基因转录本RNA编辑. 武汉植物学研究, 2010, 28: 251-256WeiL, Fei ZJ, DingY. Mitochondrial RNA editing of ATPase atp6 gene transcripts of Yunnan purple rice( Oryza sativa L. ). J Wuhan Bot Res, 2010, 28: 251-256 (in Chinese with English abstract)[本文引用:1]
[14]
刘海军, 赵丽梅, 董英山, 石瑛, 张春宝. 大豆细胞质雄性不育系及其保持系线粒体基因的RNA编辑位点研究. 分子植物育种, 2014, 12: 694-700Liu HJ, Zhao LM, Dong YS, ShiY, Zhang CB. RNA editing analysis of mitochondrial gene in cytoplasmic male sterile line and maintainer line in soybean. Mol Plant Breed, 2014, 12: 694-700 (in Chinese with English abstract)[本文引用:1]
[15]
施真, 程大友, 罗成飞, 刘巧红, 史淑芝, 崔杰. 甜菜线粒体 atp6基因转录本的RNA编辑位点研究. 分子植物育种, 2012, 10: 428-432ShiZ, Cheng DY, Luo CF, Liu QH, Shi SZ, CuiJ. The study of RNA editing sites of the atp6 gene transcripts in sugar-beet mitochondria. Mol Plant Breed, 2012, 10: 428-432 (in Chinese with English abstract)[本文引用:1]
JiangW, Yang SP, Yu DY, Gai JY. A comparative study of ATPase subunit 9 ( Atp9) gene between cytoplasmic male sterile line and its maintainer line in soybeans. African J Biotechnol, 2011, 10: 10387-10392[本文引用:1]
MourasA, Suharsono, HernouldM, Zabaleta E, Araya A. Inheritance of an induced male-sterile trait in transgenic plants expressing an engineered unedited atp9 mitochondrial gene. Theor Appl Genet, 1999, 98: 614-621[本文引用:2]
[20]
HernouldM, SuharsonoS, ZabaletaE, Carde PJ, LitvakS, ArayaA, MourasA. Impairment of tapetum and mitochondria in engineered male-sterile tobacco plants. Plant Mol Biol, 1998, 36: 499-508[本文引用:2]
[21]
NakajimaY, YamamotoT, MuranakaT, OedaK. A novel orfB-related gene of carrot mitochondrial genomes that is associated with homeotic cytoplasmic male sterility (CMS). Plant Mol Biol, 2001, 46: 99-107[本文引用:1]
[22]
DasS, SenS, ChakrabortyA, ChakrabortiP, Maiti MK, BasuA, BasuD, Sen SK. An unedited 1. 1 kb mitochondrial orfB gene transcript in the wild abortive cytoplasmic male sterility (WA-CMS) system of Oryza sativaL. subsp. indica. BMC Plant Biol, 2010, 10: 76-78[本文引用:1]
[23]
袁清华, 谢锐鸿, 张振臣, 马柱文, 李集勤, 李淑玲, 陈俊标. 烟草表达抗病基因同源物(RGA)的鉴定及RGA-SSR标记的开发. 作物学报, 2014, 40: 240-252Yuan QH, Xie RH, Zhang ZC, Ma ZW, Li JQ, Li SL, Chen JB. Identification of expressed resistance gene analogues (RGAs) and development of RGA-SSR markers in Nicotiana. Acta Agron Sin, 2014, 40: 240-252 (in Chinese with English abstract)[本文引用:1]
[24]
WangR, ZengZ, LiuT, LiuA, ZhaoY, LiK, ChenL. A novel formaldehyde metabolic pathway plays an important role during formaldehyde metabolism and detoxification in tobacco leaves under liquid formaldehyde stress. Plant Physiol Biochem, 2016, 105: 233-241[本文引用:1]
[25]
陈荣平, 刘磊, 万秀清, 邱恩建, 王春军, 宋宝刚, 颜培强, 杨铁钊. TMV侵染烟草基因差异表达的cDNA-AFLP分析. 作物学报, 2012, 38: 62-70Chen RP, LiuL, Wan XQ, Qiu EJ, Wang CJ, Song BG, Yan PQ, Yang T Z. cDNA-AFLP analysis of differentially expressed genes in tobacco infected by TMV. Acta Agron Sin, 2012, 38: 62-70 (in Chinese with English abstract)[本文引用:1]
[26]
刘齐元, 朱滕义, 赵婷, 黄海泉. 异常线粒体基因 ORF25可能导致烟草雄性不育. 江西农业大学学报, 2009, 31: 54-57Liu QY, Zhu TY, ZhaoT, Huang HQ. Aberrant mitochondrial gene ORF25may cause tobacco male sterility. Acta Agric Univ Jiangxiensis, 2009, 31: 54-57 (in Chinese with English abstract)[本文引用:1]
[27]
周玮, 刘齐元, 陈雪峰, 刘飞虎, 曹槐, 刘次全. 与烟草细胞质雄性不育相关的线粒体基因 atp9的mRNA研究. 广西植物, 2007, 27: 114-120ZhouW, Liu QY, Chen XF, Liu FH, CaoH, Liu CQ. Studies on atp9 mRNA related to cytoplasmic male sterility in tobacco. Guihaia, 2007, 27: 114-120 (in Chinese with English abstract)[本文引用:1]
周玮, 黄河, 周伯楠, 刘齐元. 烟草CMS相关基因 orf25的生物信息学分析. 中国农学通报, 2011, 27(5): 312-316ZhouW, HuangH, Zhou BN, Liu QY. The bioinformatics analysis of orf25 gene related to tobacco cytoplasmic male sterility. Chin Agric Sci Bull, 2011, 27(5): 312-316 (in Chinese with English abstract)[本文引用:1]
徐航, 刘仁祥, 徐如宏, 聂琼, 段兴友, 陈绍凯, 王国琴, 袁富, 蔡琳. 烟草RNA 提取方法的优化及冰盒储存时间的探讨. 江西农业大学学报, 2014, 36: 495-500XuH, Liu RX, Xu RH, NieQ, Duan XY, Chen SK, Wang GQ, YuanF, CaiL. The optimization of RNA isolation and the exploration of ice box storage time in tobacco. Acta Agric Univ Jiangxiensis, 2014, 36: 495-500 (in Chinese with English abstract)[本文引用:1]
[33]
杨锦华. 茎用芥菜细胞质雄性不育相关基因的克隆及机制研究. 浙江大学博士学位论文, 浙江杭州, 2006Yang JH. Characterization and molecular mechanism of cytoplasmic male-sterility associated genes in stem mustard ( Brassica juncea var. tumidaTsen et Lee). PhD Dissertation of Zhejiang University, Hangzhou, China, 2006 (in Chinese with English abstract)[本文引用:2]
[34]
AliK, TyagiA. Recent advances in acetyl CoA carboxylase: a key enzyme of fatty acid biosynthesis in plants. Intl J Appl Biol Pharmac Technol, 2016, 7: 264-277[本文引用:1]
[35]
LichtK, Jantsch MF. Rapid and dynamic transcriptome regulation by RNA editing and RNA modifications. J Cell Biol, 2016, 213: 15-22[本文引用:1]
[36]
ArayaA, DomecC, BeguD, LitvakS. An in vitro system for the editing of ATP synthase subunit 9 mRNA using wheat mitochondrial extracts. Proc Natl Acad Sci USA, 1992, 89: 1040-1044[本文引用:1]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}