摘要以综合性状优良的烤烟种质Y3为轮回亲本, 抗烟草黑胫病(0号和1号生理小种)和赤星病的雪茄烟种质Beinhart 1000-1及优质烤烟品种K326为供体亲本, 连续回交并结合分子标记辅助选择, 构建国内外第一套由256个具有烤烟Y3遗传背景并渐渗有Beinhart 1000-1和K326染色体片段株系代换系群体。该群体携带377个代换片段, 分布于烟草24条连锁群上。每个株系携带1~5个代换片段, 代换片段长度介于0.05~36.88 cM, 平均长度7.75 cM。代换片段重叠累加总长度为2922.57 cM, 是烟草基因组总长度的2.61倍。代换片段覆盖总长度为1114.32 cM, 烟草基因组覆盖率为99.45%。本研究构建的片段代换系可用于烟草基因定位、复杂性状的QTL分析和标记辅助选择育种。
关键词:烟草( Nicotiana tabacumL.); 染色体片段代换系; SSR; 分子标记辅助选择 Construction and Genetic Evaluation of Chromosome Segment Substitution Lines in Tobacco ( Nicotiana tabacum L.) TONG Zhi-Jun, JIAO Fang-Chan, FANG Dun-Huang, CHEN Xue-Jun, WU Xing-Fu, ZENG Jian-Min, XIE He, ZHANG Yi-Han, XIAO Bing-Guang* Yunnan Academy of Tobacco Agricultural Sciences / Key Laboratory of Tobacco Biotechnological Breeding / National Tobacco Genetic Engineering Research Center, Kunming 650021, China Fund:This study was supported by China National Tobacco Company [2014TBB01, 110201301006(JY-06)] and Yunnan Tobacco Company (2014YN18, 2013YN01) AbstractA set of chromosome segment substitution lines (CSSLs) of tobacco ( Nicotiana tabacum L.) were first developed by molecular marker assisted selection (MAS) and successive backcrossing with Y3, the flue-cured tobacco germplasm with comprehensive traits as the recipient parent, and two common tobacco cultivars Beinhart1000-1 and K326 as the donor parents. The cigar tobacco cultivar Beinhart 1000-1 carries a variety of resistance traits including black shank (race 0 and race 1) and brown spot resistance, while the flue-cured tobacco K326 is a commercial cultivar with high quality. In 256 CSSLs, a total of 377 substituted segments derived from donor Beinhart 1000-1 and K326 with genetic background of Y3 distributed on 24 linkage groups. Each CSSL contained only 1-5 substituted segments and the length of substituted segments ranged from 0.05 to 36.88 cM with an average of 7.75 cM. The total length of the overlapped substituted segments was 2922.57 cM, which was 2.61 times of the whole tobacco genome length. And the CSSLs covered length was 1114.32 cM, with a covered ratio of 99.45% in the recurrent tobacco genome. The CSSLs constructed in this study are excellent genetic materials for gene mapping, QTL analysis of complicated traits, and developing varieties by marker assisted selection in Nicotiana tabacum L.
Keyword:Tobacco ( Nicotiana tabacum L.); Chromosome segment substitution lines (CSSLs); SSR; Molecular marker assisted selection (MAS) Show Figures Show Figures
图1 烟草CSSLs群体构建程序Fig. 1 Procedure for developing CSSLs in tobacco
表2 Table 2 表2(Table 2)
表2 受体与供体亲本间的烟草SSR标记多态性 Table 2 Polymorphism between the recipient and donors detected by tobacco SSR markers
供体 Donor
检测标记数目1) No. of marker1)
多态标记数目 No. of polymorphic marker
多态率 Ratio of polymorphism (%)
多态标记间平均距离2) Average distance between polymorphic marker2) (cM)
共有多态性标记数目 No. of shared polymorphic marker
K326
18 764
626
3.34
1.79 (1120.45/626)
416
Beinhart 1000-1
18 764
562
3.00
2.39 (1341.18/562)
1)The 5119 PT-series SSR markers reported by Bindler et al. (2011) and 13 645 TM-series SSR markers (the data was unpublished) were used to screen the polymorphism between the recipient and donors; 2) calculated based on the length of the two tobacco genetic linkage maps of flue-cured tobacco (YK and YB ) map derived from two crosses Y3/K326 and Y3/Beinhart 1000-1, respectively; the data of the map related are unpublished. 1)检测标记数目由两部分组成, 5119对PT系列SSR标记由Bindler等(2011)公布, 13 645对TM系列SSR标记由本实验室开发(数据尚未公开); 2)根据本实验室利用新开发的烟草SSR标记构建的两张图谱各自长度计算(信息尚未公开)。
表2 受体与供体亲本间的烟草SSR标记多态性 Table 2 Polymorphism between the recipient and donors detected by tobacco SSR markers
图2(1) 256份代换系的代换片段在烟草基因组上的位置 黑色部分代表供体亲本K326的染色体片段; 绿色部分代表供体亲本Beinhart 1000-1的染色体片段。Fig. 2(1) Genomic positions of the substituted segment in 256 CSSLs Black bar represents the chromosomal segments of K326; Green bar represents the chromosomal segments of Beinhart 1000-1.
图2(2) 256份代换系的代换片段在烟草基因组上的位置 黑色部分代表供体亲本K326的染色体片段; 绿色部分代表供体亲本Beinhart 1000-1的染色体片段。Fig. 2(2) Genomic positions of the substituted segment in 256 CSSLs Black bar represents the chromosomal segments of K326; Green bar represents the chromosomal segments of Beinhart 1000-1.
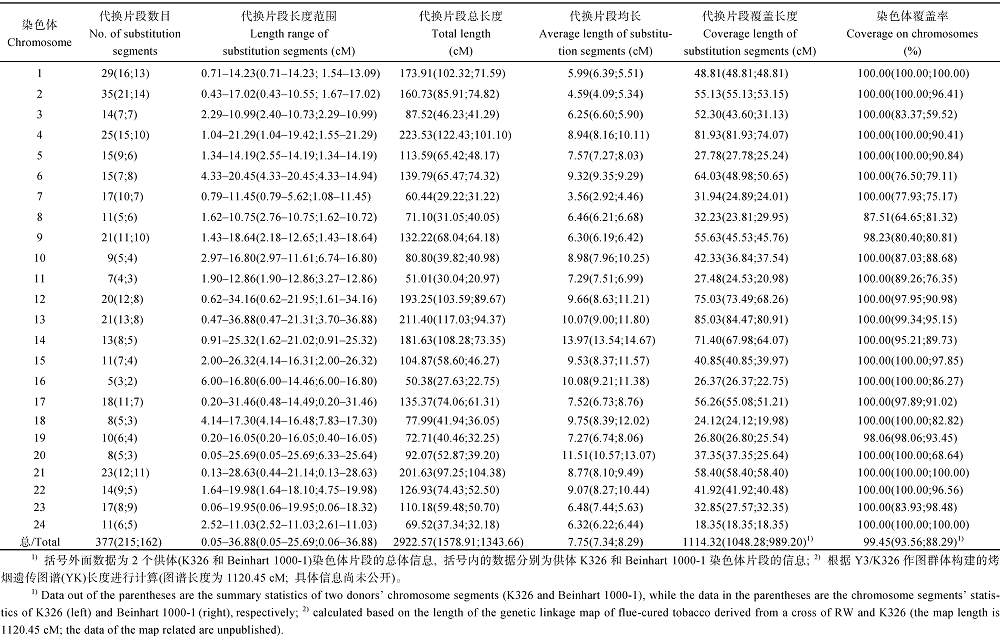
从表3和图2可看出, 烟草基因组导入的377个代换片段总长度为2922.57 cM, 长度范围为0.05~ 36.88 cM, 平均长度为7.75 cM, 最长的代换片段位于第13连锁群上, 最短的代换片段位于第20连锁群上的中下部。其中, 第4连锁群代换片段的总长最大, 为223.53 cM; 第16连锁群代换片段的总长最小, 为50.38 cM。具有烤烟K326片段的代换系中导入的215个代换片段总长度为1578.91 cM, 长度范围为0.05~25.69 cM, 平均长度为7.34 cM。具有雪茄烟Beinhart 1000-1片段的代换系中导入的162个代换片段总长度为1343.66 cM, 长度范围为0.06~36.88 cM, 平均长度为8.29 cM。 表3 Table 3 表3(Table 3)
表3 烟草染色体片段代换系的遗传评价 Table 3 Genetic evaluation of chromosome segment substitution lines in tobacco (Nicotiana tabacum L.)
表3 烟草染色体片段代换系的遗传评价 Table 3 Genetic evaluation of chromosome segment substitution lines in tobacco (Nicotiana tabacum L.)
王元英, 周健. 中美主要烟草品种亲源分析与烟草育种. 中国烟草学报, 1995, 13(5): 11-22Wang YY, ZhouJ. Genetic analysis of the main tobacco cultivars and the tobacco breeding in USA and China. Acta Tab Sin, 1995, 13(5): 11-22 (in Chinese with English abstract)[本文引用:1]
[2]
Tong ZJ, Jiao TL, Wang FQ, Li MY, Len XD, Gao YL, Li YP, Xiao BG, Wu WR. Mapping of quantitative trait loci conferring resistance to brown spot in flue-cured tobacco (Nicotiana tabacum L. ). Plant Breed, 2012, 131: 335-339[本文引用:3]
[3]
Tong ZJ, Jiao FC, Wu XF, Wang FQ, Chen XJ, Li XY, Gao YL, Zhang YH, Xiao BG, Wu WR. Mapping of quantitative trait loci underlying six agronomic traits in flue-cured tobacco (Nicotiana tabacum L. ). Acta Agron Sin, 2012, 38: 1-9[本文引用:3]
[4]
NishiT, TajimaT, NoguchiS, AjisakaH, NegishiH. Identification of DNA markers of tobacco linked to bacterial wilt resistance. Theor Appl Genet, 2003, 106: 765-770[本文引用:2]
[5]
JulioE, Denoyes RB, Verrier J L, de Borne F D. Detection of QTLs linked to leaf and smoke properties in Nicotiana tabacum based on a study of 114 recombinant inbred lines. Mol Breed, 2006, 22: 144-166[本文引用:2]
[6]
VontimittaV, Lewis RS. Mapping of quantitative trait loci affecting resistance to Phytophthora nicotianae in tobacco (Nicotiana tabacum L. ) line Beihart-1000. Mol Breed, 2010, 58: 294-300[本文引用:2]
[7]
廖长见, 王颖姮, 潘光堂. 作物染色体导入系的构建及其应用. 分子植物育种, 2007, 5(6): 139-144Liao CJ, Wang YH, Pan GT. Construction and application of chromosome introgression lines in crops. Mol Plant Breed, 2007, 5(6): 139-144 (in Chinese with English abstract)[本文引用:2]
EshedY, Zam IR. An introgression line population of Lycopersicon pennellii in the cultivated tomato enables the identification and fine mapping of yield-associated QTL. Genetics, 1995, 14: 1147-1162[本文引用:3]
[11]
Monforte AJ, Tanksley SD. Development of a set of near isogenic and backcross recombinant inbred lines containing most of the Lycopersicon hirsutum genome in an L. esculentum genetic background: A tool for gene mapping and gene discovery. Genome, 2000, 43: 803-813[本文引用:2]
[12]
BernacehiD, Beck-BunnT, EmmattyD, EshedY, InaiS, LoPez J, Petiard V, Sayama H, U’hlig J, Zamir D, Tanksley S. Advanced backcross QTL analysis of tomato: II. Evaluation of near-isogenic lines carrying single-donor introgressions for desirable wild QTL-alleles derived from Lycopersicon hirsutum and L. pimpinellifolium. Theor Appl Genet, 1998, 97: 170-180[本文引用:2]
[13]
Ramsay LD, Jennings DE, Bohuon E J R, Arihur A E, Lydiate D J, Kearsey M J, Marshall D F. The construction of a substitution library of recombinant backcross lines in Brassica oleracea for the precision mapping of quantitative trait loci. Genome, 1996, 39: 558-567[本文引用:1]
[14]
Jeuken M JW, LindhoutP. The development of lettuce backcross inbred lines (BILs) for exploitation of the Laetuca saligna (wild lettuce) germplasm. Thero Appl Genet, 2004, 109: 394-401[本文引用:2]
[15]
MatusL, CoreyA, FiliehkinT. Development and characterization of recombinant chromosome substitution lines (RCSLs) using Hordeum velgare subsp. spontaneum as a source of donor alleles in a Hordeum vulgar subsp. velgare background. Genome, 2003, 46: 10l0-1023[本文引用:2]
[16]
Korff MV, WangH, LeonJ. Development of cand idate introgression lines using an exotic barley accession (Hordeum vulgar subsp. spontaneous) as donor. Theor Appl Genet, 2004, 109: 1736-1745[本文引用:1]
[17]
冯长辉, 张胜昔, 王志伟, 王夏青, 李夕梅. 棉花染色体单片段导入系的研究进展. 湖北农业科学, 2009, 48(12): 24-26Feng CH, Zhang SX, Wang ZW, Wang XQ, Li XM. Research progress on construction and application of SSILs in cotton. Hubei Agric Sci, 2009, 48(12): 24-26 (in Chinese with English abstract)[本文引用:1]
[18]
孟佳丽, 娄群峰, 周晓慧, 史建磊, 陈劲枫. 黄瓜-酸黄瓜染色体片段导入系群体的构建及果实相关数量性状基因的定位. 中国农业科学, 2012, 45: 1558-1567Meng JL, Lou QF, Zhou XH, Shi JL, Chen JF. Construction of cucumber-sour cucumber chromosome introgression lines and location of fruit related QTLs. Sci Agric Sin, 2012, 45: 1558-1567 (in Chinese with English abstract)[本文引用:2]
[19]
林肖剑, 许学文, 钱红梅, 齐晓花, 徐强, 陈学好. 黄瓜抗白粉病染色体片段导入系的SSR鉴定. 园艺学报, 2012, 39: 485-492Lin JX, Xu XW, Qian HM, Qi XH, XuQ, Chen XH. Analysis of cucumber chromosome segment introgression line with powdery mildew resistance based on SSR markers. Acta Hort Sin, 2012, 39: 485-492 (in Chinese with English abstract)[本文引用:1]
[20]
史建磊, 娄群峰, 钱春桃, 万红建, 周晓慧, 陈劲枫. 黄瓜染色体片段导入系的构建与遗传评价. 南京农业大学学报, 2011, 34(1): 20-24Shi JL, Lou QF, Qian CT, Wan HJ, Zhou XH, Chen JF. Construction and genetic evaluation of chromosome segment introgression lines in cucumber. J Nanjing Agric Univ, 2011, 34(1): 20-24 (in Chinese with English abstract)[本文引用:1]
[21]
刘冠明, 李文涛, 曾瑞珍, 张泽民, 张桂权. 水稻单片段代换系片段的QTL鉴定. 遗传学报, 2004, 31: 1395-1400Liu GM, Li WT, Zeng RZ, Zhang ZM, Zhang GQ. Identification of QTLs on substituted segments in single segment substitution lines of rice. Acta Genet Sin, 2004, 31: 1395-1400 (in Chinese with English abstract)[本文引用:2]
[22]
何风华, 席章营, 曾瑞珍, AkshayT, 张桂权. 利用高代回交和分子标记辅助选择建立水稻单片段代换系. 遗传学报, 2005, 32: 825-831He FH, Xi ZY, Zeng RZ, AkshayT, Zhang GQ. Developing single segment substitution lines (SSSls) in rice (Oryza sativa L. ) using advanced backcrosses and MAS. Acta Genet Sin, 2004, 32: 825-831 (in Chinese with English abstract)[本文引用:3]
[23]
Li ZK, Fu BY, Gao YN. Genome-wide introgression lines and their use in genetic and molecular dissection of complex phenotypes in rice (Oryza sativa L. ). Plant Mol Biol, 2005, 59: 33-52[本文引用:2]
[24]
王玉民, 席章营, 尚爱兰, 王帮太. 作物单片段代换系的构建及应用. 中国农学通报, 2008, 24(3): 67-71Wang YM, Xi ZY, Shang AL, Wang BT. Construction and application of single segment substitution lines in crops. Chin Agric Sci Bull, 2008, 24(3): 67-71 (in Chinese with English abstract)[本文引用:2]
[25]
Bindler, G, der Hoeven VR, GunduzI, PlieskeJ, GanalM, RossiL, GadaniF, DoniniP. A microsatellite marker based linkage map of tobacco. Theor Appl Genet, 2007, 114: 341-349[本文引用:1]
[26]
Bindler, G, PlieskeJ, BakaherN, GunduzI, Ivanov N, der Hoeven R V, Ganal M, Donini P. A high density genetic map of tobacco (Nicotiana tabacum L. ) obtained from large scale microsatellite marker development. Theor Appl Genet, 2011, 123: 219-230[本文引用:1]
[27]
Sierro, N, Battey J N D, Ouadi S, Bakaher N, Bovet L, Willig A, Goepfert S, Peitsch M C, Ivanov N V. The tobacco genome sequence and its comparison with those of tomato and potato. Nat. Commun, 2014, 5: 3833, doi: DOI:10.1038/ncomms4833[本文引用:2]
[28]
Tong ZJ, Yang ZM, Chen XJ, Jiao FC, Li XY, Wu XF, Gao YL, Xiao BG, Wu WR. Large-scale development of microsatellite markers in Nicotiana tabacum and construction of a genetic map of flue-cured tobacco. Plant Breed, 2012, 131: 674-680[本文引用:3]
[29]
ArslanB, OkunusA. Genetic and geographic polymorphism of cultivated tobaccos (Nicotiana tabacum) in Turkey. Plant Genet, 2006, 42: 667-671[本文引用:1]
[30]
Del PianoL, Abet MC, SorrentinoF, AcanforaE, CozzolinoE, DiMuro A. Genetic variability in Nicotiana tabacum and Nicotiana species as revealed by RAPD procedure. Intl Contribut Tob Res, 2000, 19: 1-15[本文引用:1]
[31]
Moon HS, Nicholson JS, Lewis RS. Use of transferable Nicotiana tabacum L. microsatellite markers for investigating genetic diversity in the genus Nicotiana. Genome, 2008, 51: 547-559[本文引用:1]
[32]
Moon HS, Nicholson JS, HeinemanA, Lion K, der Hoeven R V, Hayes A J, Lewis R S. Changes in genetic diversity of U. S. flue-cured tobacco germplasm over seven decades of cultivar development. Crop Sci, 2009, 49: 498-506[本文引用:1]
[33]
Moon HS, Nifong JM, Nicholson JS, HeinemanA, Lion K, der Hoeven R V, Hayes A J, Lewis R S. Microsatellite-based analysis of tobacco (Nicotiana tabacum L. ) genetic resources. Crop Sci, 2009, 49: 2149-2157[本文引用:1]
[34]
BaiD, ReelederR, Brand le JE. Identification of two RAPD markers tightly linked with the Nicotiana debneyi gene for resistance to black root rot of tobacco. Theor Appl Genet, 1995, 91: 1184-1189[本文引用:1]
[35]
MaguireT, CollinsG, SedgleyM. A modified CTAB DNA extraction procedure for plants belonging to the family Proteaceae. Plant Mol Biol Rep, 1994, 12: 106-109[本文引用:1]
[36]
Yong ND, Tanlalesy SD. Restriction fragment polymorphism maps and the concept of graphical genotypes. Theor Appl Genet, 1989, 77: 95-101[本文引用:1]
[37]
张桂权, 曾瑞珍, 丁效华, 张泽民, 李文涛. 水稻单片段代换系文库的构建及其在分子育种上的应用. 见: 方宣钧, 黎志康主编. 2005植物分子育种国际学术研讨会论文集. 北京: 科学出版社, 2005. pp 106-113Zhang GQ, Zeng RZ, Ding XH, Zhang ZM, Li WT. Construction of a library of single segment substitution lines for rice breeding by design. In: Fang X J, Li Z K, eds. Proceedings of the 2005 International Conference on Plant Molecular Breeding. Beijing: Science Press, 2005. pp 106-113[本文引用:1]
 , 焦芳婵, 方敦煌, 陈学军, 吴兴富, 曾建敏, 谢贺, 张谊寒, 肖炳光
, 焦芳婵, 方敦煌, 陈学军, 吴兴富, 曾建敏, 谢贺, 张谊寒, 肖炳光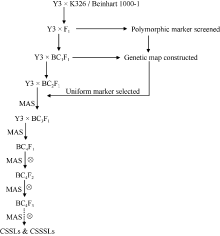





{kind=link}
{kind=link}
{kind=link}