关键词:玉米; 外源ABA; Asr1基因; 低温胁迫; 内源激素 Regulation on Contents of Endogenous Hormones and Asr1 Gene Expression of Maize Seedling by Exogenous ABA under Low-temperature Stress LI Xin-Yuan1,2,**, YANG Ye1,**, ZHANG Li-Fang1, ZUO Shi-Yu1, LI Li-Jie1, JIAO Jian1, LI Jing1,* 1 College of Agronomy, Northeast Agricultural University, Harbin 150030, China
2 Qiqihar Branch of Heilongjiang Academy of Agricultural Sciences, Qiqihaer 161006, China
Fund:This study was supported by the National Natural Science Foundation of China (31401320) AbstractAbscisic acid (ABA) is an important signal factor under low-temperature stress. This study aimed to investigate the effect of exogenous ABA on growth regulation of maize seedlings under low-temperature stress. Maize variety “Jiulong 5” resistant to low temperature was used with foliage spray of ABA (5, 15, 25, 35 mg L-1) at three-leaf period under varying degrees of low-temperature, to analyze relative electric conductivity, activities of autioxidant enzymes and contents variation of endogenous ABA and IAA of maize blades, as well as Asr1 gene expression level by Real-time PCR. The relative electric conductivity of maize blades showed an increasing tendency, meanwhile, activities of SOD and POD were enhanced by exogenous ABA treatments among which SOD activity was significantly higher in ABA treatments of 15 mg L-1 and 25 mg L-1than in the control under low-temperature stress. Also the synthesis of endogenous ABA and IAA were increased significantly. The expression level of Asr1 gene significantly promoted in ABA treatments of 5, 15, and 25 mg L-1. ABA contant had a highly significant correlation with the relative expression of Asr1 gene and SOD activity and a significant correlation with POD activity. In conclusion the expression of Asr1 gene is mediated and regulated by ABA, the promotion of Asr1 gene expression also promotes the synthesis of endogenous ABA, and enhances antioxidant enzyme activities, resulting in improved maize resistance to low temperature. But the mediation and regulation of exogenous ABA have a concentration effect, showing promotion at low concentation and inhibition at high concentation.
Keyword:Maize; Exogenous ABA; Asr1gene; Low-temperature stress; Endogenous hormone Show Figures Show Figures
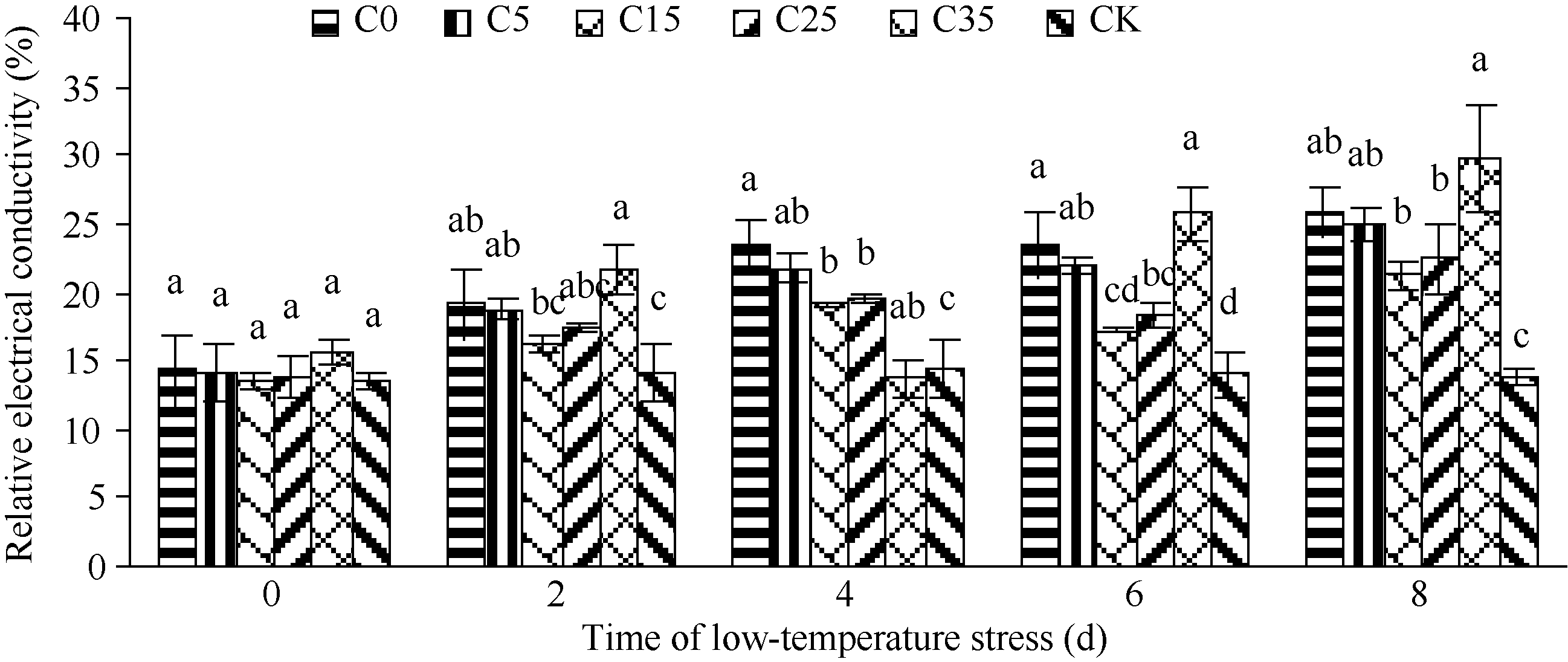
图1 外源ABA对低温胁迫下玉米苗期相对电导率的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 1 Effect of exogenous ABA treatments on cell membrane permeability in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05).
图2 外源ABA对低温胁迫下玉米苗期SOD活性的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)Fig. 2 Effect of ABA on SOD activity in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05).
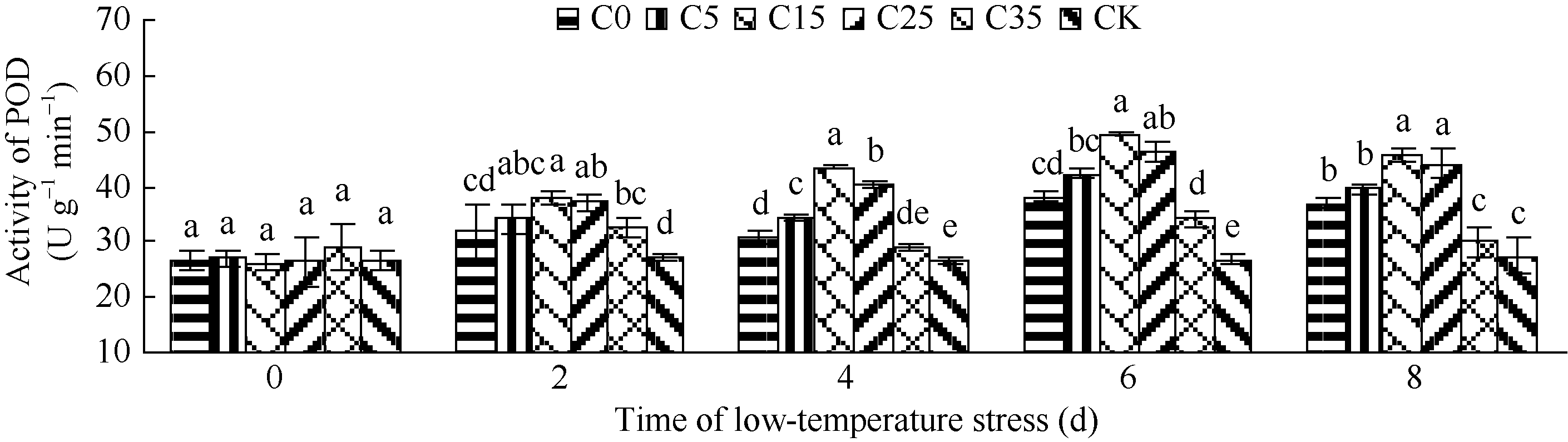
图3 外源ABA对低温胁迫下玉米苗期POD活性的影响 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。注: 柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 3 Effect of ABA on POD activity in maize seedlings under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar mean significant difference between them (P< 0.05).
表1 Table 1 表1(Table 1)
表1 外源ABA对低温胁迫下玉米苗期ABA含量的影响 Table 1 Effect of exogenous ABA treatments on ABA content in maize seedlings under low temperature stress (ng g-1 FW)
处理 Treatment
低温胁迫天数 Time of low temperature treatment
0 d
2 d
4 d
6 d
8 d
C0
77.83 e
94.36 d
94.55 e
95.04 d
100.56 e
C5
148.87 c
114.95 c
140.37 c
132.76 c
147.93 d
C15
155.91 b
155.66 a
144.33 b
150.55 b
172.63 a
C25
176.39 a
155.29 a
177.22 a
160.95 a
167.21 b
C35
122.89 d
146.65 b
132.54 d
148.05 b
152.47 c
CK
75.88 e
68.09 e
68.31 f
71.89 e
65.07 f
Data are means of three replicates. Means followed by different letters within the same column are significantly different at P < 0.05 between treatments. C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. 数据为3个重复的平均值。同列中不同字母表示处理间差异显著(P< 0.05)。C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。
表1 外源ABA对低温胁迫下玉米苗期ABA含量的影响 Table 1 Effect of exogenous ABA treatments on ABA content in maize seedlings under low temperature stress (ng g-1 FW)
表2 外源ABA对低温胁迫下玉米苗期IAA含量的影响 Table 2 Effect of exogenous ABA treatments on IAA content in maize seedlings under low temperature stress (ng g-1 FW)
处理 Treatment
低温胁迫天数 Time of low temperature treatment
0 d
2 d
4 d
6 d
8 d
C0
68.49 b
51.85 c
58.82 c
76.04 a
88.33 c
C5
35.98 d
61.36 b
59.79 b
66.59 c
70.35 e
C15
72.98 a
68.18 a
67.07 a
77.22 a
95.91 b
C25
57.79 c
50.99 c
67.70 a
66.44 c
105.37 a
C35
33.47 e
50.18 c
53.32 d
69.23 b
75.31 d
CK
58.89 c
35.22 d
38.96 e
48.03 d
61.00 f
Data are means of three replicates. Means followed by different letters within the same column are significantly different at P < 0.05 between treatments. C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. 数据为3个重复的平均值。同列中不同字母表示处理间差异显著(P< 0.05)。C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。
表2 外源ABA对低温胁迫下玉米苗期IAA含量的影响 Table 2 Effect of exogenous ABA treatments on IAA content in maize seedlings under low temperature stress (ng g-1 FW)
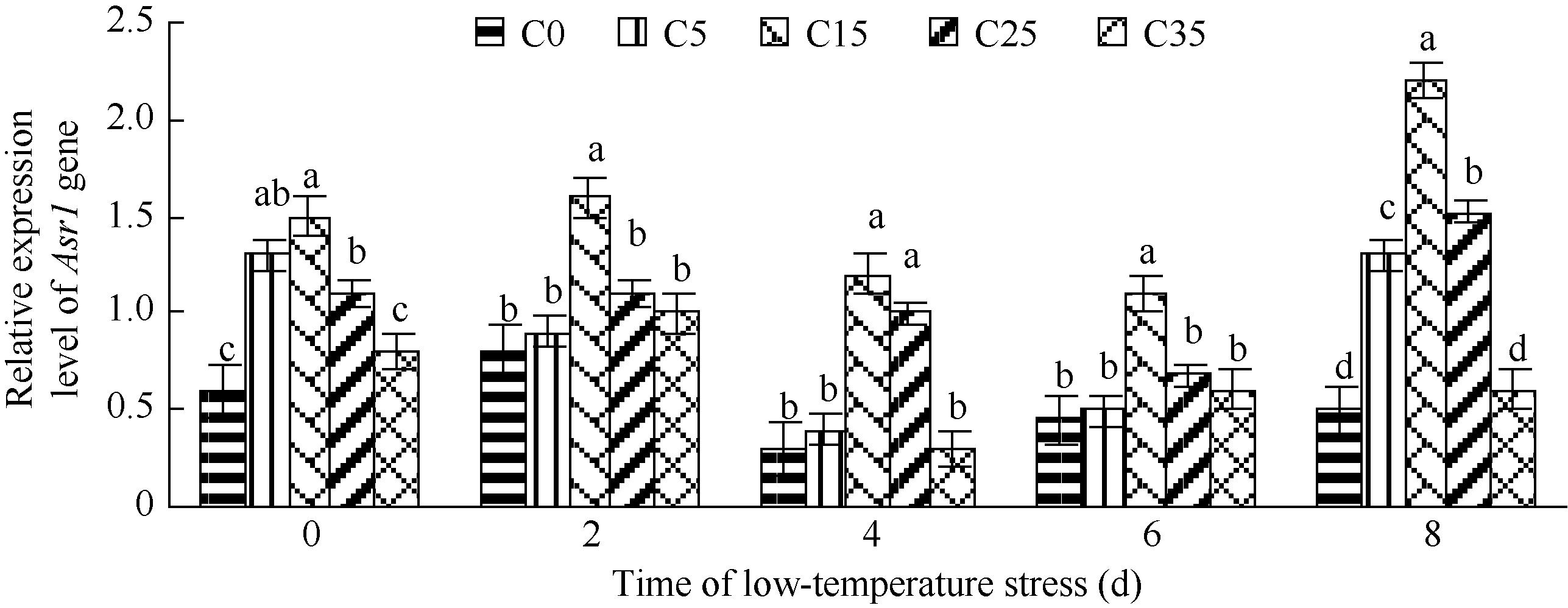
图4 低温胁迫下不同浓度外源ABA处理玉米幼苗Asr1基因相对表达量 C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。柱形图上不同字母表示处理间存在显著性差异(P< 0.05)。Fig. 4 Expression patterns of gene Asr1 in maize seedlings with different exogenous ABA treatments under low temperature stress C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. Different letters above the bar means significant difference between them (P< 0.05).
Data are means of three replicates. Means followed by different letters within the same column are significantly different at P < 0.05 between treatments. C0: treatment of water and low temperature; C5: treatment of low temperature and ABA 5 mg L-1; C15: treatment of low temperature and ABA 15 mg L-1; C25: treatment of low temperature and ABA 25 mg L-1; C35: treatment of low temperature and ABA 35 mg L-1; CK: water and room temperature. 数据为3个重复的平均值。同列中不同字母表示处理间差异显著(P< 0.05)。C0: 清水低温; C5: 低温ABA浓度5 mg L-1; C15: 低温ABA浓度15 mg L-1; C25: 低温ABA浓度25 mg L-1; C35: 低温ABA浓度35 mg L-1; CK: 清水常温。
表3 玉米幼苗干重 Table 3 Corn seedling dry weight (g)
表4 Table 4 表4(Table 4)
表4 玉米叶片激素含量、叶片干重与Asrl基因相对表达量、电导率及抗氧化酶活性的相关系数 Table 4 Correlation coefficients of leaf hormone content and leaf dry weight with Asrl gene relative expression, electrical conductivity and antioxidant enzyme activities in maize
Asrl基因相对表达量 Relative expression level of Asr1 gene
电导率 Relative electrical conductivity
SOD活性 Activity of SOD
POD活性 Activity of POD
ABA含量 ABA content
0.6939* *
0.5002
0.7094* *
0.5998*
IAA含量 IAA content
-0.9432* *
0.9187* *
0.3282
0.0322
叶片干重 Dry weight of leaf
0.7685* *
-0.5223
0.8140* *
0.6231*
* and * * indicate significant correlation at the 0.05 and 0.01 probability levels, respectively. 数值后的* 和* * 分别表示0.05和0.01水平上显著相关。
表4 玉米叶片激素含量、叶片干重与Asrl基因相对表达量、电导率及抗氧化酶活性的相关系数 Table 4 Correlation coefficients of leaf hormone content and leaf dry weight with Asrl gene relative expression, electrical conductivity and antioxidant enzyme activities in maize
潘华盛, 张桂华, 徐南平. 20世纪80年代以来黑龙江气候变暖的初步分析. 气候与环境研究, 2003, 8: 348-355Pan HS, Zhang GH, Xu NP. A preliminary analysis of climate warming in Heilongjiang province since the 1980 s. Climatic Environ Res, 2003, 8: 348-355 (in Chinese with English abstract)[本文引用:1]
[2]
史占忠, 贲显明, 张敬涛, 谷口利策, 宋光义. 三江平原春玉米低温冷害发生规律及防御措施. 黑龙江农业科学, 2003, (2): 7-10Shi ZZ, Ben XM, Zhang JT, Tanikchi, Song G Y. The emerging pattern and preventive measure of maize cold damage in the Sanjiang River Plain. Heilongjiang Agric Sci, 2003, (2): 7-10 (in Chinese with English abstract)[本文引用:1]
[3]
张建平, 王春乙, 赵艳霞, 杨晓光, 王靖. 基于作物模型的低温冷害对我国东北三省玉米产量影响. 生态学报, 2012, 32: 4132-4138Zhang JP, Wang CY, Zhao YX, Yang XG, WangJ. Impact evaluation of low temperature to yields of maize in Northeast China based on crop growth model. Acta Ecol Sin, 2012, 32: 4132-4138 (in Chinese with English abstract)[本文引用:1]
[4]
孙庆玲, 李培英, 孙宗玖, 阿不来提. 外施脱落酸对不同抗寒性狗牙根品种的渗透调节物质响应研究. 新疆农业大学学报, 2012, 35(2): 87-92Sun QL, Li PY, Sun ZJ, Abulaiti. Studies on response of spraying exogenous ABA to osmotic adjustment substance of bermudagrass varietiesa with different cold resistance. J Xinjiang Agric Univ, 2012, 35(2): 87-92 (in Chinese with English abstract)[本文引用:1]
[5]
汪月霞, 索标, 赵鹏飞, 曲小菲, 袁利刚, 赵雪娟, 赵会杰. 外源ABA对干旱胁迫下不同品种灌浆期小麦psbA基因表达的影响. 作物学报, 2011, 37: 1372-1377Wang YX, SuoB, Zhao PF, Qu XF, Yuan LG, Zhao XJ, Zhao HJ. Effect of abscisic acid treatment on psbA gene expression in two wheat cultivars during grain filling stage under drought stress. Acta Agron Sin, 2011, 37: 1372-1377 (in Chinese with English abstract)[本文引用:1]
[6]
李长宁, Srivastava MK, 农倩, 李杨瑞. 水分胁迫下外源ABA提高甘蔗抗旱性的作用机制. 作物学报, 2010, 36: 863-870Li CN, Srivastava MK, NongQ, Li YR. Mechanism of tolerance to drought in sugarcane plant enhanced by foliage dressing of abscisic acid under water stress. Acta Agron Sin, 2010, 36: 863-870 (in Chinese with English abstract)[本文引用:1]
[7]
Wang GJ, MiaoW, Wang JY, Ma DR, Li JQ, Chen WF. Effects of exogenous abscisic acid on antioxidant system in weedy and cultivated rice with different chilling sensitivity under chilling stress. J Agron Crop Sci, 2012, 199: 200-208[本文引用:1]
[8]
Bravo LA, Zúñiga GE, AlberdiM, Corcuera LJ. The role of ABA in freezing tolerance and cold acclimation in barley. Physiol Plant, 1998, 103: 17-23[本文引用:1]
[9]
黄杏, 陈明辉, 杨丽涛, 张保青, 李杨瑞. 低温胁迫下外源ABA对甘蔗幼苗抗寒性及内源激素的影响. 华中农业大学学报, 2013, 32(4): 6-11HangX, Chen MH, Yang LT, Zhang BQ, Li YR. Exogenous ABA of sugarcane seedling under low temperature stress hardiness and the effect of endogenous hormones. J Huazhong Agric Univ, 2013, 32(4): 6-11 (in Chinese with English abstract)[本文引用:1]
[10]
杨东清, 王振林, 倪英丽, 尹燕枰, 蔡铁, 杨卫兵, 彭佃亮, 崔正勇, 江文文. 高温和外源ABA对不同持绿型小麦品种籽粒发育及内源激素含量的影响. 中国农业科学, 2014, 47: 2109-2125Yang DQ, Wang ZL, Ni YL, Yin YP, CaiT, Yang WB, Peng DL, Cui ZY, Jiang WW. Effect of high temperature stress and spraying exogenous ABA post-anthesis on grain filling and grain yield in different types of stay-green wheat. Sci Agric Sin, 2014, 47: 2109-2125 (in Chinese with English abstract)[本文引用:1]
[11]
杨猛, 庄文锋, 魏湜, 顾万荣, 杨晔, 王萌, 李晶. 玉米苗期受低温胁迫蛋白表达差异研究. 核农学报, 2013, 27: 1742-1748YangM, Zhang WF, WeiS, Gu WR, YangY, WangM, LiJ. Differentially expressed proteome under low temperature stress in maize seedling. Acta Agric Nucl Sin, 2013, 27: 1742-1748 (in Chinese with English abstract)[本文引用:1]
[12]
Carpentier SC, VertomraenA, SwennenR, WittersE, FortesC, Souza MT, PanB. Sugar-mediated acclimation: the importance of sucrose metabolism in meristems. J Proteome Res, 2010, 9: 5038-5046[本文引用:1]
[13]
JeanneauM, GerentesD, FoueillassarcX, ZivydM, VidalaJ, Toppand A, PerezP. Improvement of drought tolerance in maize: towards the functional validation of the Zm-Asr1 gene and increase of water use efficiency by over-expressing C4-PEPC. Biochimie, 2002, 84: 1127-1135[本文引用:1]
[14]
Isabelle MH, Sebastien CC, SuzanP, Anais VH, PanisB, SwennenR, RemyS. Structure and regulation of the Asr gene family in banana. Planta, 2011, 234: 785-798[本文引用:1]
[15]
ShenG, Pang YZ, Wu WS, Deng ZX, Liu XF, LinJ, Zhao LX, Sun XF, Tang KX. Molecular cloning, characterization and expression of a novel Asr gene from Ginkgo biloba. Plant Physiol Biochem, 2005, 43: 836-843[本文引用:1]
[16]
FrankelN, CarrariF, HassonE. Evolutionary history of the Asr gene family. Gene, 2006, 37(8): 74-83[本文引用:1]
[17]
Chin YY, Yu CC, Guang YJ, Wang CS. A lily ASR protein involves abscisic acid signaling and confers drought and salt resistance in Arabidopsis. Plant Physiol, 2005, 139: 836-846[本文引用:1]
张新友, 徐静, 汤丰收, 董文召, 臧秀旺, 张忠信. 花生种间杂种胚胎发育及内源激素变化. 作物学报, 2013, 39(6): 1127-1133Zhang XY, XuJ, Tang FS, Dong WZ, Zang XW, Zhang ZX. Embryonic development and changes of endogenous hormones in interspecific hybrids between peanut (A. hypogaea L. ) and wild arachis species. Acta Agron Sin, 2013, 39: 1127-1133 (in Chinese with English abstract)[本文引用:1]
[20]
江福英, 李延, 翁伯琦. 植物低温胁迫及其抗性生理. 福建农业学报, 2002, 17(3): 190-195Jiang FY, LiY, Weng BQ. Review on physiology of chilling stress and chilling resistance of plants. Fujian J Agric Sci, 2002, 17(3): 190-195 (in Chinese with English abstract)[本文引用:1]
[21]
王玮, 张枫, 李德全. 外源ABA对渗透胁迫下玉米幼苗根系渗透调节的影响. 作物学报, 2002, 28: 121-126WangW, ZhangF, Li DQ. The effects of exogenous ABA on osmotic adjustment in maize roots under osmotic stress. Acta Agron Sin, 2002, 28: 121-126 (in Chinese with English abstract)[本文引用:1]
[22]
杨卫兵, 王振林, 尹燕枰, 李文阳, 李勇, 陈晓光, 王平, 陈二影, 郭俊祥, 蔡铁, 倪英丽. 外源ABA和GA对小麦籽粒内源激素含量及其灌浆进程的影响. 中国农业科学, 2011, 44: 2673-2682Yang WB, Wang ZL, Yin YP, Li WY, LiY, Chen XG, WangP, Chen EY, Guo JX, CaiT, Ni YL. Effects of spraying exogenous ABA or GA on the endogenous hormones concentration and filling of wheat grains. Sci Agric Sin, 2011, 44: 2673-2682 (in Chinese with English abstract)[本文引用:1]
[23]
ZhangY, Jiang WJ, Yu HJ. Exogenous abscisic acid alleviates low temperature-induced oxidative damage in seedlings of Cucumis sativus L. Trans CSAE, 2012, 28: 221-228[本文引用:1]
[24]
郑莎莎, 孙传范, 孙红春, 刘连涛, 赵金峰, 李存东. 不同外源激素对花铃期棉花主茎叶生理特性的影响. 中国农业科学, 2009, 42: 4383-4389Zheng SS, Sun CF, Sun HC, Liu LT, Zhao JF, Li CD. Effects of different exogenous hormones on physiological characteristics of main stem leaves at flower and boll stage in cotton. Sci Agric Sin, 2009, 42: 4383-4389 (in Chinese with English abstract)[本文引用:1]
[25]
IqbalM, AshrafM, RehmanS, Rha ES. Does polyamine seed pretreatment modulate growth and levels of some plant growth regulators in hexaploid wheat (Triticum aestivum L. ) plants under salt stress?Bot Stud, 2006, 47: 239-250[本文引用:2]
[26]
Gomez CA, ArbonaV, JacasJ, Primo ME, TalonM. Abscisic acid reduces leaf abscission and increases salt tolerance in citrus plants. Plant Growth Regul, 2003, 21: 234-240[本文引用:1]
Huang JC, Lin SM, Wang CS. A pollen-specific and desiccation associated transcript in Lilium longiflorum during development and stress. Plant Cell Physiol, 2000, 41: 477-485[本文引用:1]
[29]
程维舜, 孙玉宏, 曾红霞, 杜念华, 施先锋, 蔡新忠. ASR蛋白与植物的抗逆性研究进展. 园艺学报, 2013, 40: 2049-2057Cheng WS, Sun YH, Zeng HX, Du NH, Shi XF, Cai XZ. ASR protein and plant stress tolerance. Acta Hortic Sin, 2013, 40: 2049-2057 (in Chinese with English abstract)[本文引用:1]
[30]
Cortés AJ, Chavarro MC, MadriñánS, ThisD, Blair MW. Molecular ecology and selection in the drought related Asr gene polymorphisms in wild and cultivated common bean (Phaseolus vulgaris L. ). BMC Genet, 2012, 13: 58[本文引用:1]
[31]
GoldgurY, RomS, Ghirland oR, ShkolnikD, ShadrinN, KonradZ, Bar-ZviD. Desiccation and zinc binding induce transition of tomato abscisic acid stress ripening 1, a water stress and salt stress-regulated plant-specific protein, from unfolded to folded state. Plant Physiol, 2007, 143: 617-628[本文引用:1]
[32]
MaskinL, MaldonadoS, Iusem ND. Tomato leaf spatial expression of stress-induced Asr genes. Mol Biol Rep, 2008, 35: 501-505[本文引用:1]
[33]
VeeraragavanP, Dilip M A L. Expression analysis of a gene family in loblolly pine (Pinus taeda L. ) induced by water deficit stress. Plant Mol Biol, 1997, 35: 801-807[本文引用:1]
[34]
杨晔, 李晶, 顾万荣, 魏湜. Asr基因家族的研究进展. 作物杂志, 2013, (3): 7-11YangY, LiJ, Gu WR, WeiS. Advances in the research of Asr gene family. Crops, 2013, (3): 7-11 (in Chinese with English abstract)[本文引用:1]
 , 杨晔
, 杨晔





{kind=link}
{kind=link}
{kind=link}
{kind=link}