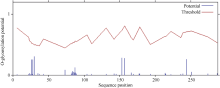
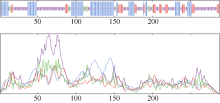
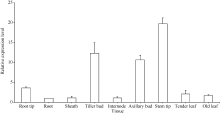
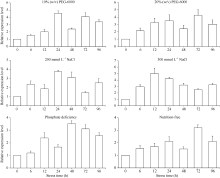
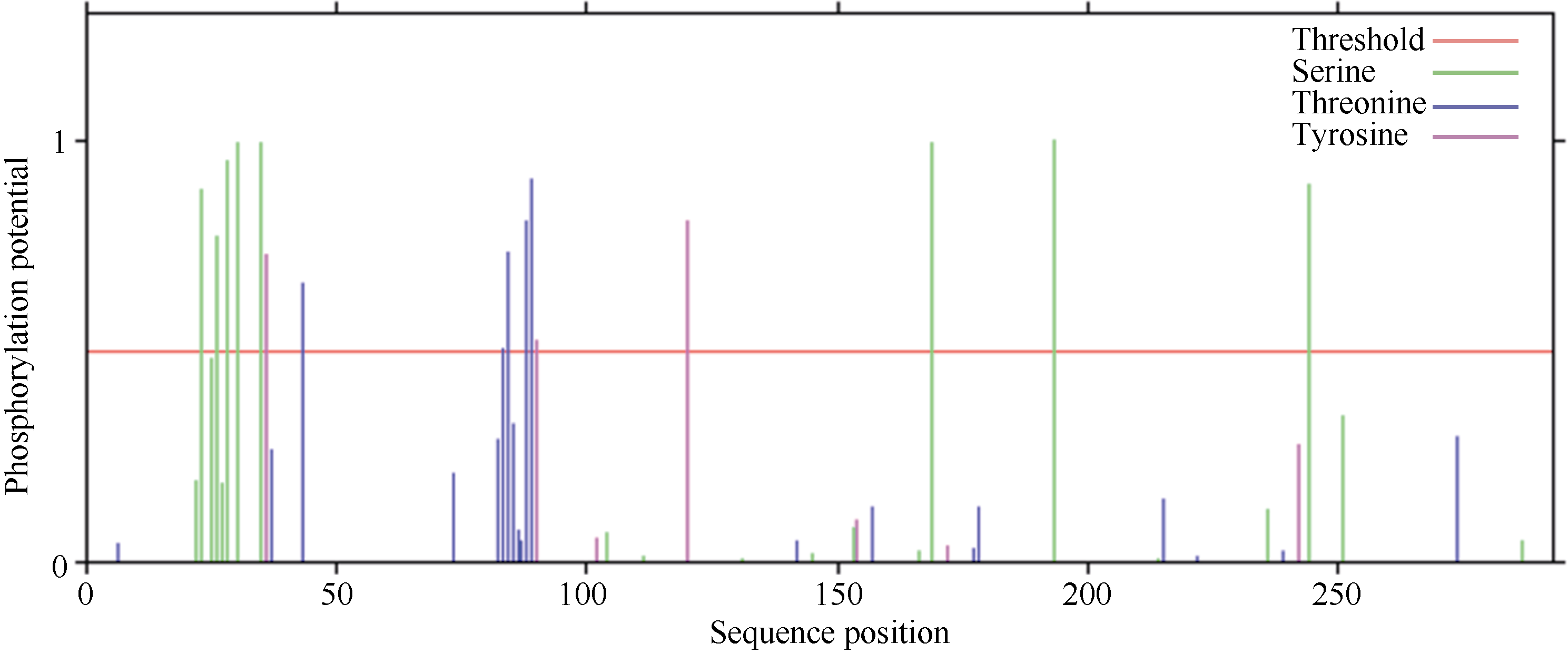
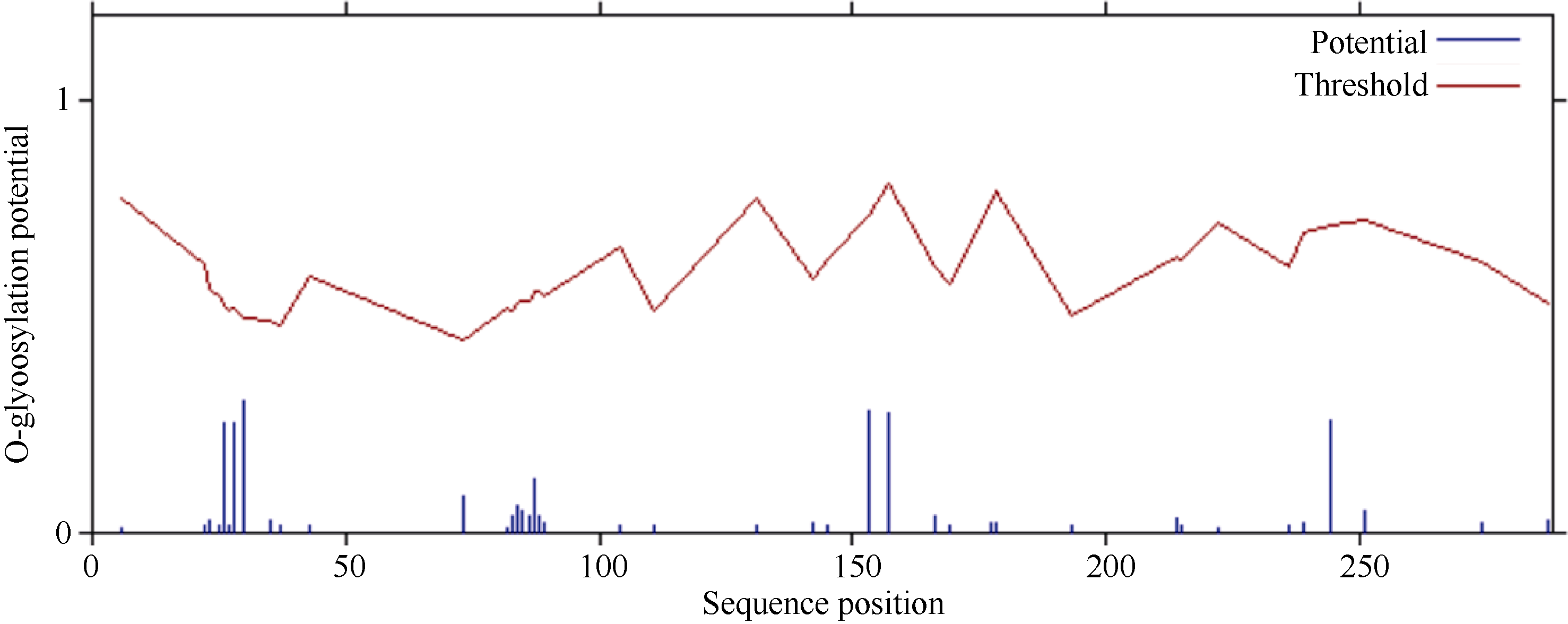
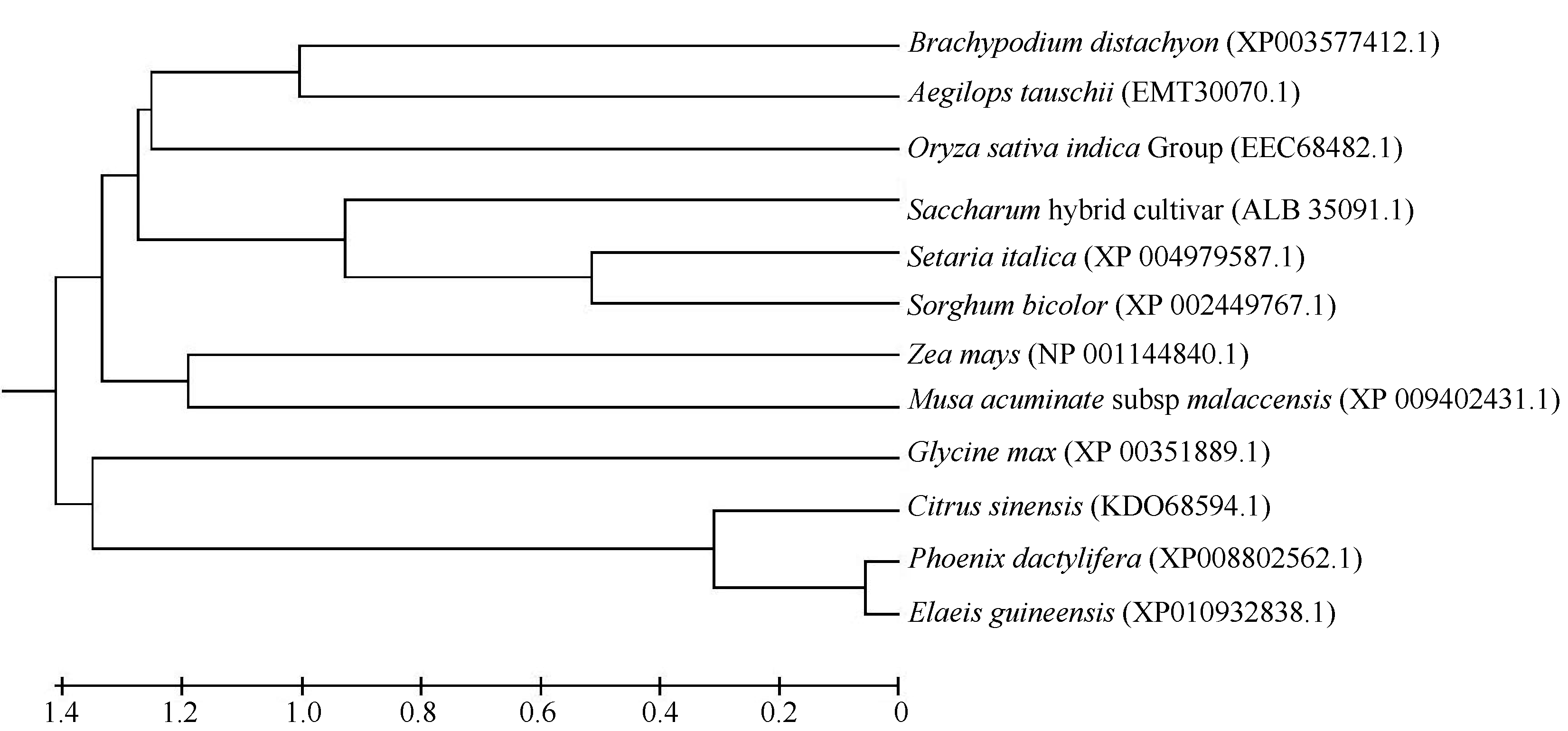
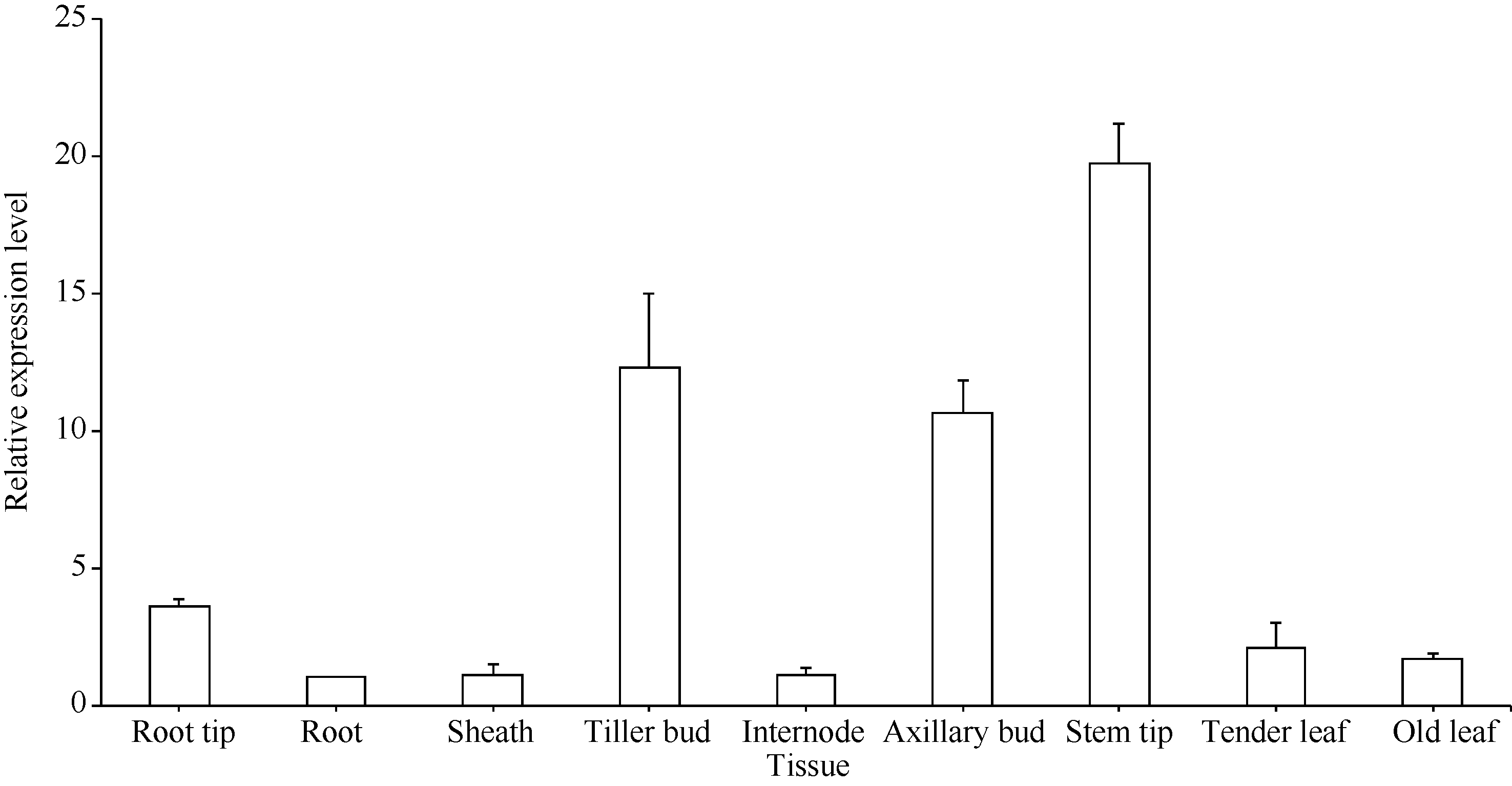
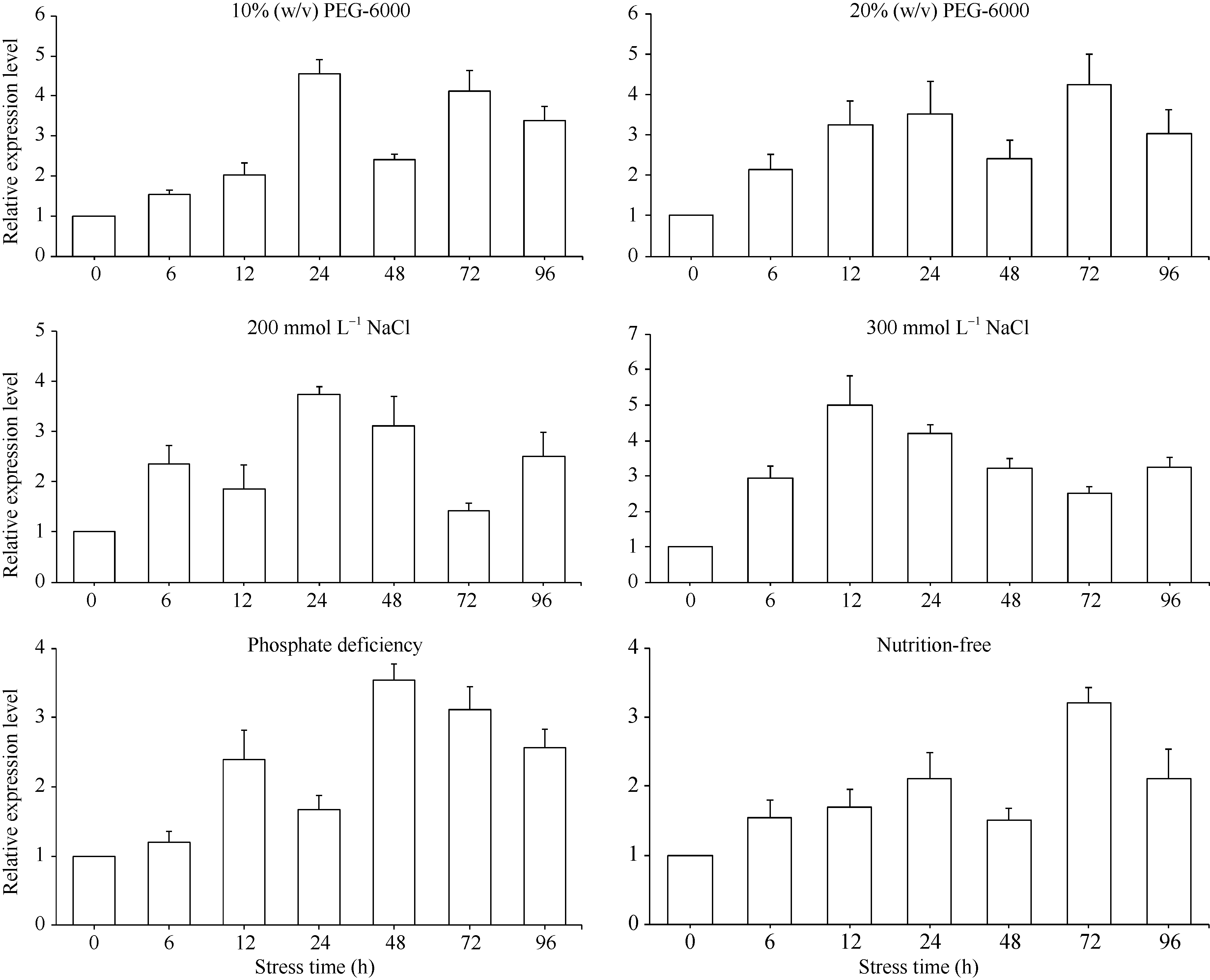
关键词:甘蔗; ScD27; 同源克隆; 生物信息学; q-PCR Cloning and Expression Analysis of Key Gene ScD27in Strigolactones Biosynthesis Pathway WU Zhuan-Di**, LIU Xin-Long**, LIU Jia-Yong, ZAN Feng-Gang, LI Xu-Juan, LIU Hong-Bo, LIN Xiu-Qin, CHEN Xue-Kuan, SU Huo-Sheng, ZHAO Pei-Fang, WU Cai-Wen* Sugarcane Research Institute, Yunnan Academy of Agricultural Sciences / Yunnan Key Laboratory of Sugarcane Genetic Improvement, Kaiyuan 661699, China Fund:This study was supported by the National Natural Science Foundation of China (31360359), the National Modern Agricultural Industry Technology System Construction Project (CARS-20-1-1), the Young and Middle-aged Academic Technology Leaders Reserve Talented Person in Yunnan Province (2014HB038), the Applied Basic Research Projects in Yunnan Province (2016FB071), the Major Science and Technology Projects - Biology (2015ZA001), and the Science and Technology Innovation Talents Project (2014HC015) AbstractStrigolactones (SLs) is a novel class of plant hormones. D27 regulating reversible metabolic process is located in up-stream of strigolactones biosynthesis pathway. In this study, primers were designed based on the conserved domains from four species inluding Oryza sativa, Zea mays, Sorghum bicolor, and Brachypodium distachyon. Using cDNA from sugarcane cultivar ROC22 as the template, the full-length cDNA sequence of D27gene from sugarcane was cloned by RT-PCR and RACE method. This gene is named as ScD27, with the GenBank accession number of KP987221.1. Its length is 1379 bp, and it contains an 867 bp open reading frame (ORF), encoding 288 amino acid residues. ScD27 is not a secretory protein and has a molecular weight of 71.58 kD, with a theoretical isoelectric point of 5.04. ScD27 is mainly located in chloroplast and the conserved domains of this protein involve two zinc finger protein structures (ZnF_TAZ and ZnF_A20). Amino acid sequences encoded by ScD27shared more than 70% similarity with the reported amino acid sequences encoded by D27 of Sorghum bicolor, Setaria italicaBeauv., Hordeum vulgare subsp. vulgare and Brachypodium distachyon. ScD27 gene was differentially expressed in different parts of sugarcane plant, with higher level of transcript in stem tip and axillary bud but much lower level in leaf, stem and root. Furthermore, the expression of ScD27 could be induced by the stresses of PEG, salt and the deficiencies of phosphorus and nutrition. These results demonstrated that ScD27 might be a key gene participating in the response to abiotic stresses during sugarcane SLs biosynthesis pathway.
许智宏, 李家洋. 中国植物激素研究: 过去、现在和未来. 植物学通报, 2006, 23: 433-442Xu ZH, Li JY. Plant hormones research in china: past, present and future. Chin Bull Bot, 2006, 23: 433-442 (in Chinese with English abstract)[本文引用:1]
[2]
王威豪, 王一丁, 莫云川, 叶燕萍, 李杨瑞. 水分胁迫下喷施乙烯利对甘蔗分蘖及农艺性状的影响. 广西农业科学, 2007, (2): 148-151Wang WH, Wang YD, Mo YC, Ye YP, Li YY. Studies on effects of etheph on physiological, biochemical and agronomical characters in sugarcane during tillering stage under water stress. Guangxi Agric Sci, 2007, (2): 148-151 (in Chinese with English abstract)[本文引用:1]
[3]
VasanthaS, Shekinah DE, GuptaC, RakkiyappanP. Tiller production, regulation and senescence in sugarcane (Saccharum species hybrid) genotypes. Sugar Tech, 2012, 14: 156-160[本文引用:1]
[4]
Gomez-RoldanV, FermasS, Brewer PB, Puech-PagesV, Dun EA, Pillot JP, LetisseF, MatusovaR, DanounS, Portais JC, BouwmeesterH, BecardG, Beveridge CA, RameauC, Rochange SF. Strigolactone inhibition of shoot branching. Nature, 2008, 455: 189-194[本文引用:1]
UmeharaM, HanadaA, YoshidaS, AkiyamaK, AriteT, Takeda-KamiyaN, MagomeH, KamiyaY, ShirasuK, YoneyamaK, KyozukaJ, YamaguchiS. Inhibition of shoot branching by new terpenoid plant hormones. Nature, 2008, 455: 195-200[本文引用:2]
[7]
KoltaiH. Strigolactones are regulators of root development. New Phytol, 2011, 190: 545-549[本文引用:1]
[8]
Al-BabiliS, Bouwmeester HJ. Strigolactones, a novel carotenoid-derived plant hormone. Annu Rev Plant Biol, 2015, 66: 161-186[本文引用:1]
[9]
RasmussenA, Hosseini SA, Hajirezaei MR, DruegeU, GeelenD. Adventitious rooting declines with the vegetative to reproductive switch and involves a changed auxin homeostasis. J Exp Bot, 2015, 66: 1437-1452[本文引用:1]
[10]
Andreo-JimenezB, Ruyter-SpiraC, Bouwmeester HJ, Lopez- Raez J A. Ecological relevance of strigolactones in nutrient uptake and other abiotic stresses, and in plant-microbe interactions below-ground. Plant Soil, 2015, 394: 1-19[本文引用:1]
[11]
AlderA, JamilM, MarzoratiM, BrunoM, VermathenM, BiglerP, GhislaS, BouwmeesterH, BeyerP, Al-BabiliS. The path from beta-carotene to carlactone, a strigolactone-like plant hormone. Science, 2012, 335: 1348-1351[本文引用:1]
[12]
BookerJ, SiebererT, WrightW, WilliamsonL, WillettB, StirnbergP, TurnbullC, SrinivasanM, GoddardP, LeyserO. MAX1 encodes a cytochrome P450 family member that acts downstream of MAX3/4 to produce a carotenoid-derived branch-inhibiting hormone. Dev Cell, 2005, 8: 443-449[本文引用:1]
[13]
Simons JL, Napoli CA, Janssen BJ, Plummer KM, Snowden KC. Analysis of the DECREASED APICAL DOMINANCE genes of petunia in the control of axillary branching. Plant Physiol, 2007, 143: 697-706[本文引用:1]
[14]
Drummond RS, Martinez-Sanchez N M, Janssen B J, Templeton K R, Simons J L, Quinn B D, Karunairetnam S, Snowden K C. Petunia hybrida CAROTENOID CLEAVAGE DIOXYGENASE 7 is involved in the production of negative and positive branching signals in petunia. Plant Physiol, 2009, 151: 1867-1877[本文引用:1]
[15]
Drummond RS, SheehanH, Simons JL, Martinez-Sanchez N M, Turner R M, Putterill J, Snowden K C. The expression of petunia strigolactone pathway genes is altered as part of the endogenous developmental program. Front Plant Sci, 2011, 2: 115[本文引用:1]
[16]
SorefanK, BookerJ, HaurogneK, GoussotM, BainbridgeK, FooE, ChatfieldS, WardS, BeveridgeC, RameauC, LeyserO. MAX4 and RMS1 are orthologous dioxygenase-like genes that regulate shoot branching in Arabidopsis and pea. Genes Dev, 2003, 17: 1469-1474[本文引用:1]
[17]
王涛. 水稻独脚金内酯相关基因的图位克隆与功能分析. 中国农业科学院博士学位论文, 北京, 2012WangT. Map-Based Cloning and Functional Analysis of the Strigolactones-Related Genes in Rice (Orzya sativa L. ). PhD Dissertation of Chinese Academy of Agriculture Science, Beijing, China, 2012 (in Chinese with English abstract)[本文引用:2]
[18]
Waters MT, Brewer PB, Bussell JD, Smith SM, Beveridge CA. The Arabidopsis ortholog of rice DWARF27 acts upstream of MAX1 in the control of plant development by strigolactones. Plant Physiol, 2012, 159: 1073-1085[本文引用:3]
[19]
ZhangY, van Dijk AD, ScaffidiA, Flematti GR, HofmannM, CharnikhovaT, VerstappenF, HepworthJ, van der KrolS, LeyserO, Smith SM, ZwanenburgB, Al-BabiliS, Ruyter-SpiraC, Bouwmeester HJ. Rice cytochrome P450 MAX1 homologs catalyze distinct steps in strigolactone biosynthesis. Nat Chem Biol, 2014, 10: 1028-1033[本文引用:1]
[20]
LinH, WangR, QianQ, YanM, MengX, FuZ, YanC, JiangB, SuZ, LiJ, WangY. DWARF27, an iron-containing protein required for the biosynthesis of strigolactones, regulates rice tiller bud outgrowth. Plant Cell, 2009, 21: 1512-1525[本文引用:6]
[21]
Rubio-MoragaA, AhrazemO, Perez-Clemente R M, Gomez-Cadenas A, Yoneyama K, Lopez-Raez J A, Molina R V, Gomez-Gomez L. Apical dominance in saffron and the involvement of the branching enzymes CCD7 and CCD8 in the control of bud sprouting. BMC Plant Biol, 2014, 14: 171[本文引用:1]
[22]
陈芳育, 江良荣, 郑景生, 黄荣裕, 王侯聪. 用蛋白质组遗传方法分析水稻多蘖矮秆突变体的共分离蛋白. 中国遗传学会大会, 厦门大学, 福建厦门, 2011Chen FY, Jiang LR, Zheng JS, Huang RY, Wang HC. Fine mapping and proteomics analysis of a high-tillering dwarf mutant in rice. Congress of Chinese Genetic Society, Xiamen University, Xiamen, China, 2010[本文引用:1]
Harrison PJ, Newgas SA, DescombesF, Shepherd SA, Thompson AJ, Bugg TD. Biochemical characterization and selective inhibition of beta-carotene cis-trans isomerase D27 and carotenoid cleavage dioxygenase CCD8 on the strigolactone biosynthetic pathway. FEBS J, 2015, 282: 3986-4000[本文引用:1]
[25]
WenC, ZhaoQ, NieJ, LiuG, ShenL, ChengC, XiL, MaN, ZhaoL. Physiological controls of chrysanthemum DgD27 gene expression in regulation of shoot branching. Plant Cell Rep, 2016[本文引用:2]
[26]
AbeS, SadoA, TanakaK, KisugiT, AsamiK, OtaS, Kim HI, YoneyamaK, XieX, OhnishiT, SetoY, YamaguchiS, AkiyamaK, YoneyamaK, NomuraT. Carlactone is converted to carlactonoic acid by MAX1 in Arabidopsis and its methyl ester can directly interact with AtD14 in vitro. Proc Natl Acad Sci USA, 2014, 111: 18084-18089[本文引用:2]
[27]
ZouJ, ZhangS, ZhangW, LiG, ChenZ, ZhaiW, ZhaoX, PanX, XieQ, ZhuL. The rice HIGH-TILLERING DWARF1 encoding an ortholog of Arabidopsis MAX3 is required for negative regulation of the outgrowth of axillary buds. Plant J, 2006, 48: 687-698[本文引用:1]
[28]
王闵霞, 白玉路, 王平, 向跃武, 蔡平钟, 张志雄. 水稻Dwarf 14(D14)基因的生物信息学分析. 西南农业学报. 2014, 27: 1347-1352Wang MX, Bai YL, WangP, Xiang YW, Cai PZ, Zhang ZX. Bioinformatics analysis of D14 (dwarf 14) in rice. Southwest China J Agric Sci, 2014, 27: 1347-1352 (in Chinese with English abstract)[本文引用:1]
[29]
ZhaoJ, WangT, WangM, LiuY, YuanS, GaoY, YinL, SunW, PengL, ZhangW, WanJ, LiX. DWARF3 participates in an SCF complex and associates with DWARF14 to suppress rice shoot branching. Plant Cell Physiol, 2014, 55: 1096-1109[本文引用:1]
[30]
Zhao LH, Zhou XE, YiW, WuZ, LiuY, KangY, Hou L, de Waal P W, Li S, Jiang Y, Scaffidi A, Flematti G R, Smith S M, Lam V Q, Griffin P R, Wang Y, Li J, Melcher K, Xu H E. Destabilization of strigolactone receptor DWARF14 by binding of ligand and E3-ligase signaling effector DWARF3. Cell Res, 2015, 25: 1219-1236[本文引用:1]
[31]
SoundappanI, BennettT, MorffyN, LiangY, Stanga JP, AbbasA, LeyserO, Nelson DC. SMAX1-LIKE/D53 family members enable distinct MAX2-dependent responses to strigolactones and karrikins in Arabidopsis. Plant Cell, 2015, 27: 3143-3159[本文引用:1]
[32]
AriteT, UmeharaM, IshikawaS, HanadaA, MaekawaM, YamaguchiS, Kyozuka J. d14, a strigolactone-insensitive mutant of rice, shows an accelerated outgrowth of tillers. Plant Cell Physiol, 2009, 50: 1416-1424[本文引用:1]
[33]
IshikawaS, MaekawaM, AriteT, OnishiK, TakamureI, KyozukaJ. Suppression of tiller bud activity in tillering dwarf mutants of rice. Plant Cell Physiol, 2005, 46: 79-86[本文引用:1]
JiangL, LiuX, XiongG, LiuH, ChenF, WangL, MengX, LiuG, YuH, YuanY, YiW, ZhaoL, MaH, HeY, WuZ, MelcherK, QianQ, Xu HE, WangY, LiJ. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature, 2013, 504: 401-405[本文引用:1]
[36]
Brewer PB, Dun EA, Ferguson BJ, RameauC, Beveridge CA. Strigolactone acts downstream of auxin to regulate bud outgrowth in pea and Arabidopsis. Plant Physiol, 2009, 150: 482-493[本文引用:1]
[37]
贾昆鹏. 植物激素独脚金内酯和茉莉酸信号与光信号互作的分子机制研究. 上海交通大学博士学位论文, 上海, 2014Jia KP. The Molecular Mechanism of Cross Talking between Light and Phytohormones Strigolactone and Jasmonate Signaling. PhD Dissertation of Shanghai Jiao Tong University, Shanghai, China, 2014 (in Chinese with English abstract)[本文引用:2]
[38]
KumarM, Pand ya-KumarN, DamA, HaorH, Mayzlish-GatiE, BelausovE, WiningerS, Abu-AbiedM, McErlean C S, Bromhead L J, Prand i C, Kapulnik Y, Koltai H. Arabidopsis response to low-phosphate conditions includes active changes in actin filaments and PIN2 polarization and is dependent on strigolactone signalling. J Exp Bot, 2015, 66: 1499-1510[本文引用:1]
[39]
YoneyamaK, XieX, KusumotoD, SekimotoH, SugimotoY, TakeuchiY, YoneyamaK. Nitrogen deficiency as well as phosphorus deficiency in sorghum promotes the production and exudation of 5-deoxystrigol, the host recognition signal for arbuscular mycorrhizal fungi and root parasites. Planta, 2007, 227: 125-132[本文引用:1]
[40]
Lopez-Raez JA, CharnikhovaT, Gomez-RoldanV, MatusovaR, KohlenW, De VosR, VerstappenF, Puech-PagesV, BecardG, MulderP, BouwmeesterH. Tomato strigolactones are derived from carotenoids and their biosynthesis is promoted by phosphate starvation. New Phytol, 2008, 178: 863-874[本文引用:1]
[41]
BuQ, LvT, ShenH, LuongP, WangJ, WangZ, HuangZ, XiaoL, EngineerC, Kim TH, Schroeder JI, HuqE. Regulation of drought tolerance by the F-box protein MAX2 in Arabidopsis. Plant Physiol, 2014, 164: 424-439[本文引用:1]
[42]
Ha CV, Leyva-Gonzalez M A, Osakabe Y, Tran U T, Nishiyama R, Watanabe Y, Tanaka M, Seki M, Yamaguchi S, Dong N V, Yamaguchi-Shinozaki K, Shinozaki K, Herrera-Estrella L, Tran L S. Positive regulatory role of strigolactone in plant responses to drought and salt stress. Proc Natl Acad Sci USA, 2014, 111: 851-856[本文引用:1]
[43]
YamaguchiS, KyozukaJ. Branching hormone is busy both underground and overground. Plant Cell Physiol, 2010, 51: 1091-1094[本文引用:1]
[44]
SunH, TaoJ, LiuS, HuangS, ChenS, XieX, YoneyamaK, ZhangY, XuG. Strigolactones are involved in phosphate- and nitrate-deficiency-induced root development and auxin transport in rice. J Exp Bot, 2014, 65: 6735-6746[本文引用:2]
[45]
李晓君. 甘蔗逆境相关锌指蛋白基因ShSAP1的功能研究. 海南大学博士学位论文, 海南海口, 2012Li XJ. Functional Analysis of Stress Associated Zinc Finger Protein Gene ShSAP1 from Sugarcane. PhD Dissertation of Hainan University, Haikou, China, 2012 (in Chinese with English abstract)[本文引用:1]
[46]
阙友雄, 许莉萍, 徐景升, 张积森, 张木清, 陈如凯. 甘蔗基因表达定量PCR分析中内参基因的选择. 热带作物学报, 2009, 30(3): 274-278Que YX, Xu LP, Xu JS, Zhang JS, Zhang MQ, Chen RK. Selection of control genes in Real-time qPCR analysis of gene expression in sugarcane. Chin J Trop Crops, 2009, 30(3): 274-278 (in Chinese with English abstract)[本文引用:1]
[47]
谢晓娜, 杨丽涛, 王盛, 张小秋, 李杨瑞. 甘蔗NADP异柠檬酸脱氢酶基因(SoNADP-IDH)的克隆与表达分析. 中国农业科学, 2015, 48: 185-196Xie XN, Yang LT, WangS, Zhang XQ, Li YY. Cloning and expression analysis of sugarcane NADP+-dependent isocitrate dehydrogenase (SoNADP-IDH) gene. Sci Agric Sin, 2015, 48: 185-196 (in Chinese with English abstract)[本文引用:1]
[48]
VijS, Tyagi AK. Genome-wide analysis of the stress associated protein (SAP) gene family containing A20/AN1 zinc-finger(s) in rice and their phylogenetic relationship with Arabidopsis. Mol Genet Genom, 2006, 276: 565-575[本文引用:1]
[49]
WaldieT, MccullochH, LeyserO. Strigolactones and the control of plant development: lessons from shoot branching. Plant J, 2014, 79: 607-622[本文引用:1]
[50]
李晓君, 武媛丽, 孔冉, 杨本鹏, 张树珍. 植物A20/AN1型锌指蛋白基因功能研究进展. 生物技术通报, 2013, 29(12): 6-14Li XJ, Wu YL, KongR, Yang BP, Zhang SZ. Functional Research of A20/AN1 type zinc finger protein gene in plants. Biotechnol Bull, 2013, 29(12): 6-14 (in Chinese with English abstract)[本文引用:1]
[51]
Mayzlish-GatiE, De-CuyperC, GoormachtigS, BeeckmanT, VuylstekeM, Brewer PB, Beveridge CA, YermiyahuU, KaplanY, EnzerY, WiningerS, ResnickN, CohenM, KapulnikY, KoltaiH. Strigolactones are involved in root response to low phosphate conditions in Arabidopsis. Plant Physiol, 2012, 160: 1329-1341[本文引用:1]
[52]
YoneyamaK, YoneyamaK, TakeuchiY, SekimotoH. Phosphorus deficiency in red clover promotes exudation of orobanchol, the signal for mycorrhizal symbionts and germination stimulant for root parasites. Planta, 2007, 225: 1031-1038[本文引用:1]
 , 刘新龙
, 刘新龙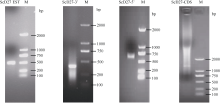











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}