关键词:小麦; 抗白粉病基因; 分子标记; 混池ddRAD测序 Development of Markers Closely Linked with Wheat Powdery Mildew Resistance Gene Pm48 FU Bi-Sheng1,**, LIU Ying1,2,**, ZHANG Qiao-Feng1, WU Xiao-You1, GAO Hai-Dong3, CAI Shi-Bin1, DAI Ting-Bo2,*, WU Ji-Zhong1,* 1 Institute of Food Crops, Jiangsu Academy of Agricultural Sciences / Jiangsu Provincial Platform for Conservation and Utilization of Agricultural Germplasm, Nanjing 210014, China
2 College of Agriculture, Nanjing Agricultural University, Nanjing 210095, China
3 Genepioneer Biotechnologies Co. Ltd., Nanjing 210014, China
Fund:This study was supported by the National Key Technology R&D Program of China (2013BAD01B02-12), the China Agriculture Research System (CARS-3-1-17), Jiangsu Provincial Foundation of Agricultural Scientific Innovation [CX (14)5006], and the Natural Science Foundation of Jiangsu Province (BK2012783) Abstract Pm48 is a novel powdery mildew resistance gene identified previously in our laboratory. This study aimed at developing close molecular markers for fine mapping of the gene. The ddRAD-sequencing assay revealed 81 SNPs associated with the target gene, in which one converted into the STS marker Xmp931 and three converted into the CAPS markers Xmp928, Xmp930, and Xmp936. We also developed 71 genomic SSR markers according to the genome sequence of Aegilops tauschii. And mapped two of them, Xmp1089 and Xmp1112. Using the 115 F2:3 families derived from the cross of Ningnuomai 1 × Tabasco, the target gene was found to be co-segregated with Xmp928 and distal to Xmp1112 with the genetic distance of 3.1 cM towards centromere. In the 671 homozygous susceptible families, Xmp928 also showed co-segregated with the target gene. We also physically mapped Pm48 to the bin of 5DS 0.63-0.67 by using three Chinese Spring 5DS deletion lines.
Keyword:Wheat; Powdery mildew resistance gene; Molecular markers; Bulked ddRAD-seq Show Figures Show Figures
图1Xmp928(A)、Xmp930(B)和Xmp1089(C)在两亲本及抗、感池的扩增带型 M: 50 bp DNA ladder; 1: Tabasco; 2: 宁糯麦1号; 3: 抗病池; 4: 感病池。Fig. 1 Polymorphism patterns between the parents and the bulks detected with markers Xmp928(A)、Xmp930(B), and Xmp1089(C) M: 50 bp DNA ladder; 1: Tabasco; 2: Ningnuomai 1; 3: resistant pool; 4: susceptible pool.
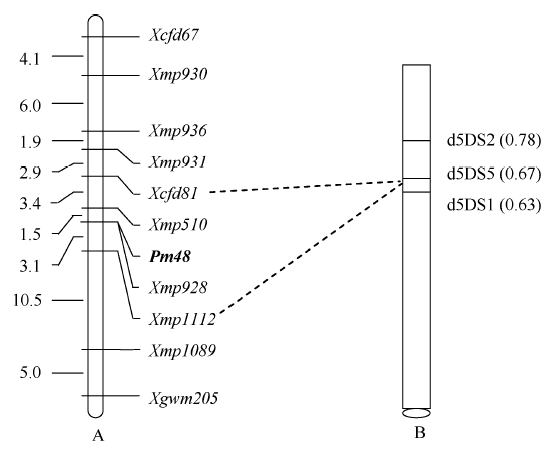
图2 抗白粉病基因Pm48的遗传图谱(A)和物理图谱(B) A图中左侧数字为遗传距离(cM); B图中右侧括号内数字是着丝粒到染色体断点处长度与染色体短臂总长度的比值, 下方椭圆代表着丝粒。Fig. 2 Genetic (A) and physical (B) maps of the powdery mildew resistance gene Pm48 In linkage map A, numbers on the left indicate genetic distances (cM). In physical map B, numbers in the parentheses indicate the ratio of length from centromere to chromosome breakpoint/total length of the short arm of chromosome and the ellipse at the bottom shows the position of centromere.
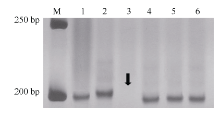
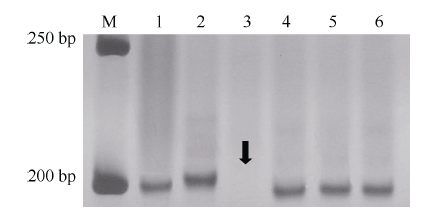
图3Xmp1112在杂交亲本和中国春5DS缺体中的扩增带型 M: 50 bp DNA ladder; 1: Tabasco; 2: 宁糯麦1号; 3~5: 中国春缺体系del5DS-1、del5DS-2和del5DS-5; 6: 中国春。箭头指示目标带缺失。Fig. 3 Profiles of Xmp1112 amplified in cross parents and Chinese Spring 5DS deletion lines M: 50 bp DNA ladder; 1: Tabasco; 2: Ningnuomai 1; 3-5: Chinese Spring deletion lines del5DS-1, del5DS-2 and del5DS-5; 6: Chinese Spring. The arrow shows vacancy of target band.
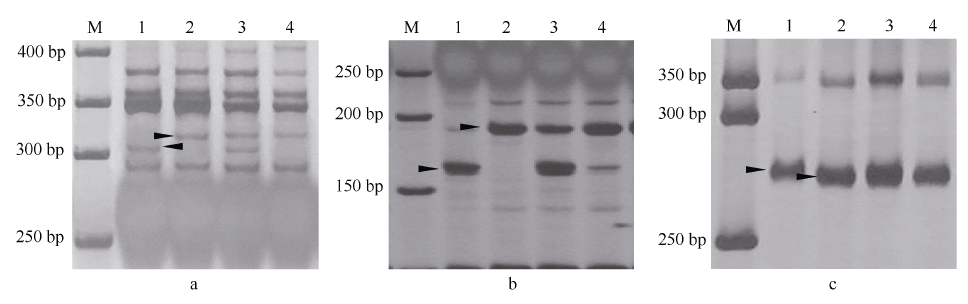
表2 19个纯合感病重组体基因型分析和分类 Table 2 Genotype classification of the 19 homozygous susceptible recombinants
重组体类型 Recombinant type
重组体单株数 No. of recombinant
重组体基因型Genotype of recombinant
Xmp1112
Xmp928
Xmp510
Xcfd81
I
11
AB
BB
BB
BB
II
1
AB
BB
A_
AB
III
4
BB
BB
A_
AB
IV
3
BB
BB
BB
AB
Only one recombination occurred in types I, III, and IV at the marker interval of Xmp1112-Xmp928, Xmp928-Xmp510, and Xmp510- Xcfd81, respectively; whereas twice recombinations occurred in type II at marker intervals of Xmp1112-Xmp928 and Xmp928-Xmp510. A: Tabasco allele; B: Ningnuomai 1 allele; A_: dominant genotype including Tabasco genotype and heterozygous genotype. 重组体I、III、IV只发生1次重组, 分别在Xmp1112-Xmp928、Xmp928-Xmp510、Xmp510-Xcfd81标记区间; 重组体II发生2次重组, 分别在Xmp1112-Xmp928和Xmp928-Xmp510标记区间。A: Tabasco等位位点; B: 宁糯麦1号等位位点; A_: 显性基因型, 包括Tabasco基因型和杂合基因型。
表2 19个纯合感病重组体基因型分析和分类 Table 2 Genotype classification of the 19 homozygous susceptible recombinants
JohnsonJ, BaenzigerP, YamazakiW, SmithR. Effects of powdery mildew on yield and quality of isogenic lines of ‘Chancellor’ wheat. Crop Sci, 1979, 19: 349-352[本文引用:1]
[2]
XiaoM, SongF, JiaoJ, WangX, XuH, LiH. Identification of the gene Pm47 on chromosome 7BS conferring resistance to powdery mildew in the Chinese wheat land race Hongyanglazi. Theor Appl Genet, 2013, 126: 1397-1403[本文引用:1]
[3]
MohlerV, BauerC, SchweizerG, KempfH, HartlL. Pm50: a new powdery mildew resistance gene in common wheat derived from cultivated emmer. J Appl Genet, 2013, 54: 259-263[本文引用:1]
[4]
ZhanH, LiG, ZhangX, LiX, GuoH, GongW, JiaJ, QiaoL, RenY, YangZ, ChangZ. Chromosomal location and comparative genomics analysis of powdery mildew resistance gene Pm51 in a putative wheat-Thinopyrum ponticum introgression line. PLoS One, 2014, 9: e113455[本文引用:1]
[5]
XuH, YiY, MaP, QieY, FuX, XuY, ZhangX, AnD. Molecular tagging of a new broad-spectrum powdery mildew resistance allele Pm2c in Chinese wheat land race Niaomai. Theor Appl Genet, 2015, 128: 2077-2084[本文引用:1]
[6]
HytenD, CannonS, SongQ, WeeksN, FickusE, ShoemakerR, SpechtJ, FarmerA, MayG, CreganP. Highthroughput SNP discovery through deep resequencing of a reduced representation library to anchor and orient scaffolds in the soybean whole genome sequence. BMC Genomics, 2010, 11: 38[本文引用:1]
[7]
PfenderW, SahaM, JohnsonE, SlabaughM. Mapping with RAD (restriction-site associated DNA) markers to rapidly identify QTL for stem rust resistance in Lolium perenne. Theor Appl Genet, 2011, 122: 1467-1480[本文引用:1]
[8]
WangN, FangL, XinH, WangL, LiS. Construction of a high-density genetic map for grape using next generation restriction-site associated DNA sequencing. BMC Plant Biol, 2012, 12: 148[本文引用:1]
[9]
Peterson BK, Weber JN, Kay EH, Fisher HS, Hoekstra HE. Double digest RADseq: an inexpensive method for de novo SNP discovery and genotyping in model and non-model species. PLoS One, 2012, 7: e37135[本文引用:1]
[10]
DavikJ, Sargent DJ, Brurberg MB, LienS, KentM, AlsheikhM. A ddRAD based linkage map of the cultivated strawberry, Fragaria xananassa. PLoS One, 2015, 10: e0137746[本文引用:1]
[11]
ZhouX, XiaY, RenX, ChenY, HuangL, HuangS, LiaoB, LeiY, YanL, JiangH. Construction of a SNP-based genetic linkage map in cultivated peanut based on large scale marker development using next-generation double-digest restriction-site-associated DNA sequencing (ddRADseq). BMC Genomics, 2014, 15: 351[本文引用:1]
[12]
WuZ, WangB, ChenX, WuJ, KingG, XiaoY, LiuK. Evaluation of linkage disequilibrium pattern and association study on seed oil content in Brassica napus using ddRAD sequencing. PLoS One, 2016, 11: e0146383[本文引用:1]
[13]
FeuilletC, TravellaS, SteinN, AlbarL, NublatA, KellerB. Map-based isolation of the leaf rust disease resistance gene Lr10 from the hexaploid wheat (Triticum aestivum L. ) genome. Proc Natl Acad Sci USA, 2003, 100: 15253-15258[本文引用:1]
[14]
YanL, LoukoianovA, TranquilliG, HelgueraM, FahimaT, DubcovskyJ. Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci USA, 2003, 100: 6263-6268[本文引用:1]
KrattingerS, LagudahE, SpielmeyerW, SinghR, Huerta-EspinoJ, McFadden H, Bossolini E, Selter L, Keller B. A putative ABC transporter confers durable resistance to multiple fungal pathogens in wheat. Science, 2009, 323: 1360-1363[本文引用:1]
[17]
YahiaouiN, SrichumpaP, DudlerR, KellerB. Genome analysis at different ploidy levels allows cloning of the powdery mildew resistance gene Pm3b from hexaploid wheat. Plant J, 2004, 37: 528-538[本文引用:1]
[18]
CaoA, XingL, WangX, YangX, WangW, SunY, QianC, NiJ, ChenY, LiuD, WangX, ChenP. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc Natl Acad Sci USA, 2011, 108: 7727-7732[本文引用:1]
[19]
GaoH, ZhuF, JiangY, WuJ, YanW, ZhangQ, JacobiA, CaiS. Genetic analysis and molecular mapping of a new powdery mildew resistant gene Pm46 in common wheat. Theor Appl Genet, 2012, 125: 967-973[本文引用:8]
Endo TR, Gill BS. The deletion stocks of common wheat. J Hered, 1996, 87: 295-307[本文引用:1]
[22]
盛宝钦. 用反应型记载小麦苗期白粉病. 植物保护, 1988, (1): 14Sheng BQ. Infection reaction types against wheat powdery mildew at seedling stage. Plant Prot, 1988, (1): 14 (in Chinese)[本文引用:1]
[23]
Ma ZQ, Sorrells ME, Tanksley SD. RFLP markers linked to powdery mildew resistance genes Pm1, Pm2, Pm3, and Pm4 in wheat. Genome, 1994, 37: 871-875[本文引用:1]
[24]
Land erE, GreenP, AbrahamsonJ, BarlowA, DaleyM, LincolnS, NewburgL. Mapmaker: an interactive computer package for constructing primary genetic linkage maps of experimental and natural populations. Genomics, 1987, 1: 174-181[本文引用:1]
[25]
刘仁虎, 孟金陵. MapDraw, 在Excel中绘制遗传连锁图的宏. 遗传, 2003, 25: 317-321Liu RH, Meng JL. MapDraw: a Microsoft Excel macro for drawing genetic linkage maps based on given genetic linkage data. Hereditas (Beijing), 2003, 25: 317-321 (in Chinese with English abstract)[本文引用:1]
[26]
KosambiD. The estimation of map distances from recombination values. Ann Eugenics, 1943, 12: 172-175[本文引用:1]
[27]
You FM, WanjugiH, HuoN, Lazo GR, Luo MC, Anderson OD, DvorakJ, Gu YQ. RJPrimers: unique transposable element insertion junction identification and primer design for marker development. Nucl Acid Res, 2010, 38: 313-320[本文引用:1]
[28]
XueS, ZhangZ, LinF, KongZ, CaoY, LiC, YiH, MeiM, ZhuH, WuJ, XuH, ZhaoD, TianD, ZhangC, MaZ. A high-density intervarietal map of the wheat genome enriched with markers derived from expressed sequence tags. Theor Appl Genet, 2008, 117: 181-189[本文引用:2]
[29]
SuenagaK, KhairallahM, WilliamH, HoisingtonD. A new intervarietal linkage map and its application for quantitative trait locus analysis of “gigas” features in bread wheat. Genome, 2005, 48: 65-75[本文引用:2]
[30]
PaillardS, SchnurbuschT, WinzelerM, MessmerM, SourdilleP, AbderhaldenO, KellerB, SchachermayrG. An integrative genetic linkage map of winter wheat (Triticum aestivum L. ). Theor Appl Genet, 2003, 107: 1235-1242[本文引用:2]
[31]
AkpinarB, MagniF, YuceM, LucasS, ŠimkováH, ŠafářJ, VautrinS, BergèsH, CattonaroF, DoleželJ, BudakH. The physical map of wheat chromosome 5DS revealed gene duplications and small rearrangements. BMC Genomics, 2015, 16: 453[本文引用:1]
[32]
EraymanM, Sand huD, SidhuD, DilbirligiM, Baenziger PS, Gill KS. Demarcating the gene-rich regions of the wheat genome. Nucl Acids Res, 2004, 32: 3546-3565[本文引用:1]
 , 刘颖
, 刘颖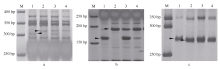


{kind=link}
{kind=link}
{kind=link}