* 通讯作者(Corresponding author): 张新友, E-mail:haasz@126.com, Tel: 0371-65729560 **同等贡献(Contributed equally to this work) 第一作者联系方式: E-mail:lilina198283@163.com, Tel: 0371-65718247 收稿日期:2016-09-18 基金:本研究由河南省重大科技专项(141100110600), 国家现代农业产业技术体系建设专项(CARS-14)和河南省现代农业产业技术体系项目(S2012-5)资助
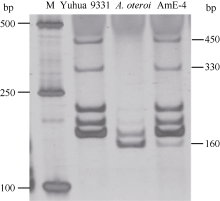
摘要花生野生种是改良花生栽培种的重要基因资源。为了利用野生花生的抗性基因, 本研究利用花生栽培品种豫花9331与二倍体野生种 A. oteroi人工杂交, 借助胚拯救和染色体秋水仙素加倍, 创制一个双二倍体杂种AmE-4, 并利用荧光原位杂交和分子标记技术准确鉴定了该双二倍体。观察结果表明, AmE-4的叶片与豫花9331存在显著差异, 而主茎高、侧枝长和总分枝数等性状与豫花9331差异不显著。AmE-4开花期较豫花9331推迟60 d, 结实性与荚果发育状况较差, 不利于AmE-4的育种利用。同时, 开发了57个追踪AmE-4中 A. oteroi染色体的显性或共显性SSR标记, 为创制和鉴定花生栽培种 A. oteroi易位系或渐渗系奠定分子基础。
关键词:花生; 双二倍体; 花生野生种; 分子标记; 荧光原位杂交 Development and Characterization of Amphidiploid Derived from Interspecific Cross between Cultivated Peanut and Its Wild Relative Arachis oteroi LI Li-Na1,2,**, DU Pei2,**, FU Liu-Yang2,3, LIU Hua2, XU Jing2, QIN Li2, YAN Mei2, HAN Suo-Yi2, HUANG Bing-Yan2, DONG Wen-Zhao2, TANG Feng-Shou2, ZHANG Xin-You2,* 1College of Agricultural, Henan University of Science and Technology, Luoyang 471023, China
2 Industrial Crops Research Institute, Henan Academy of Agricultural Sciences / Key Laboratory of Oil Crops in Huanghuaihai Plains, Ministry of Agriculture / Henan Provincial Key Laboratory for Oil Crops Improvement, Zhengzhou 450002, China
3 School of Life Sciences, Zhengzhou University, Zhengzhou 450001, China
Fund:This study was supported by the Major Technology Research and Development Program of Henan Province (141100110600), the China Agriculture Research System (CARS-14), and the Henan Provincial Agriculture Research System (S2012-05) AbstractWild Arachis species are important genetic resources. To introgress resistant genes of Arachis species, we developed a new amphidiploid AmE-4 through man-made cross between cultivated peanut variety Yuhua 9331 and a diploid Arachis species A. oteroi, with the assistance of following embryo rescue and chromosome doubling by colchicine treatment. AmE-4 was identified and characterized by fluorescence in situ hybridization (FISH) and SSR molecular marker. Morphological observation revealed significant differences in leaves between amphidiploid AmE-4 and Yuhua 9331, while the agronomic traits such as main stem height, length of first lateral branch and number of branches showed less difference between them. The date of first flower appearance in AmE-4 delayed sixty days compared with that in Yuhua 9331, and its pods setting and development were also poor, which would hinder its further utilization. In addition, 57 dominant or co-dominant SSR molecular markers were developed and could be used to identify translocation or introgression lines with A. oteroi chromosome fragment in future studies.
Keyword:Peanut ( Arachis hypogaea L.); Amphidiploid; Wild Arachis species; Molecular marker; FISH Show Figures Show Figures
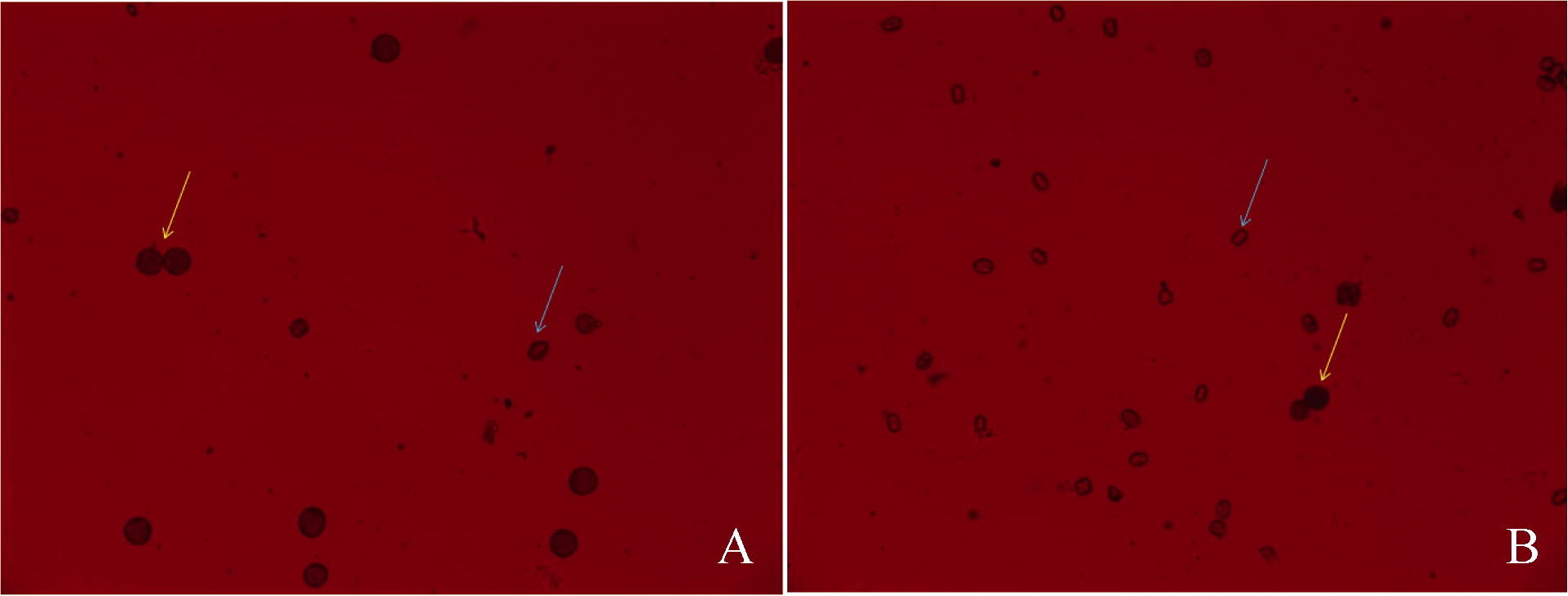
图1 S0有果针枝(A)和无果针枝(B)花的花粉染色 黄色箭头所示为可育花粉粒, 蓝色箭头所示为不育花粉粒。Fig. 1 Pollen staining from flowers in branches with (A) and without pegs (B) Yellow arrows show fertile pollens and blue arrows show sterile pollens.
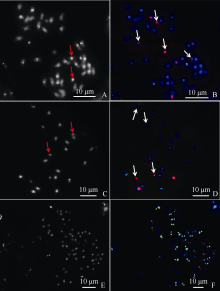
图3 豫花9331、A. oteroi和AmE-4荧光原位杂交 A~B: 豫花9331 DAPI染色及45S rDNA (红)和5S rDNA (绿)探针荧光原位杂交; C~D: A. oteroi DAPI染色及45S rDNA (红)和5S rDNA (绿)探针荧光原位杂交; E~F: AmE-4 DAPI染色和A.oteroi全基因组探针原位杂交。红色箭头所示为小染色体, 白色箭头所示为“ 大随体染色体” ; F图中的白色* 表示强基因组杂交信号的染色体, F图中的红色+表示弱基因组杂交信号的染色体。Fig. 3 Fluorescence in situ hybridization of Yuhua 9331, A. oteroi and AmE-4 A-B: DAPI staining and FISH of Yuhua9331 with 45S rDNA (red) and 5S rDNA (green) probes; C-D: DAPI staining and FISH of A. oteroi with 45S rDNA (red) and 5S rDNA (green) probes; E-F: DAPI staining and GISH of AmE-4 with A. oteroi total genomic DNA probes. Red arrows show the small chromosomes, white arrows show “ the big SAT-chromosomes” ; * show chromosomes with strong signals of A. oteroi total genomic DNA probes, and + show chromosomes with weak signals of A. oteroi total genomic DNA probes in Fig. F.
MallikarjunaN, Jadhav DR, ReddyK, HusainF, DasK. Screening new Arachis amphidiploids and autotetraploids for resistance to late leaf spot by detached leaf technique. Eur J Plant Pathol, 2012, 132: 17-21[本文引用:1]
[4]
姜慧芳, 任小平, 黄家权, 雷永, 廖伯寿. 野生花生脂肪酸组成的遗传变异及远缘杂交创造高油酸低棕榈酸花生新种质. 作物学报, 2009, 35: 25-32Jiang HF, Ren XP, Huang JQ, LeiY, Liao BS. Genetic variation of fatty acid components in Arachis species and development of interspecific hybrids with high oleic and low palmitic acids. Acta Agron Sin, 2009, 35: 25-32 (in Chinese with English abstract)[本文引用:3]
[5]
Singh AK. Hybridization barriers among the species of Arachis L. , namely of the sections Arachis (including the groundnut) and Erectoides. Genet Resour Crop Evol, 1998, 45: 41-45[本文引用:1]
[6]
张新友, 徐静, 汤丰收, 董文召, 臧秀旺, 张忠信. 花生种间杂种胚胎发育及内源激素变化. 作物学报, 2013, 39: 1127-1133Zhang XY, XuJ, Tang FS, Dong WZ, Zang XW, Zhang ZX. Embryonic development and changes of endogenous hormones in interspecific hybrids between peanut (A. hypogaea L. ) and wild Arachis species. Acta Agron Sin, 2013, 39: 1127-1133 (in Chinese with English abstract)[本文引用:1]
[7]
MallikarjunaN, SenthilvelS, HoisingtonD. Development of synthetic groundnuts (Arachis hypogaea L. ) to broaden the genetic base of cultivated groundnut. Genet Resour Crop Evol, 2011, 58: 889-907[本文引用:1]
[8]
Church GT, Starr JL, Simpson CE. A recessive gene for resistance to meloidogyne arenaria in interspecific Arachis spp. hybrids. Russ J Nematol, 2005, 37: 178-184[本文引用:1]
[9]
贺梁琼, 熊发前, 钟瑞春, 韩柱强, 李忠, 唐秀梅, 蒋菁, 唐荣华, 何新华. 利用SCoT标记分析花生栽培种 × A. chacoensis组合异源多倍化的早期基因组变化. 中国农业科学, 2013, 46: 1555-1563He LQ, Xiong FQ, Zhong RC, Han ZQ, LiZ, Tang XM, JiangJ, Tang RH, He XH. Study on genome variations by using SCoT markers during allopolyploidization of the cultivated peanut × A. chacoensis. Sci Agric Sin, 2013, 46: 1555-1563 (in Chinese with English abstract)[本文引用:1]
[10]
Ozias-AkinsP, SingsitC, Branch WD. Interspecific hybrid inviability in crosses of Arachis hypogaea × A. stenosperma can be overcome by in vitro embryo maturation or somatic embryogenesis. J Plant Physiol, 1992, 140: 207-212[本文引用:1]
Tallury SP, Isleib TG, Stalker HT. Developing peanut cultivars with genetic resistance to early leafspot. In: Proceedings of the American Peanut Research and Educational Society, 2005. p 37[本文引用:1]
[13]
王兴军, 张新友. 花生生物技术研究. 北京: 科学出版社, 2015. p 22Wang XJ, Zhang XY. Studies on Peanut Biotechnology. Beijing: Beijing Science Press, 2015. p 22[本文引用:1]
[14]
MallikarjunaN, Sastri DC. Morphological, cytological and disease resistance studies of the intersectional hybrid between Arachis hypogaea L. and A. glabrata Benth. Euphytica, 2002, 126: 161-167[本文引用:2]
[15]
MallikarjunaN, HoisingtonD. Peanut improvement: production of fertile hybrids and backcross progeny between Arachis hypogaea and A. kretschmeri. Food Secur, 2009, 1: 457-462[本文引用:1]
[16]
Stalker HT, Beute MK, Shew BB, Barker KR. Registration of two root-knot nematode-resistant peanut germplasm lines. Crop Sci, 2002, 42: 312-313[本文引用:1]
[17]
PasupuletiJ, RamaiahV, RathoreA, RupakulaA, Reddy RK, WaliyarF, Nigam SN. Genetic analysis of resistance to late leaf spot in interspecific groundnuts. Euphytica, 2013, 193: 13-25[本文引用:1]
Ferguson ME, Burow MD, Schulze SR, Bramel PJ, Paterson AH, KresovichS, MitchellS. Microsatellite identification and characterization in peanut (A. hypogaea L. ). Theor Appl Genet, 2004, 108: 1064-1070[本文引用:1]
[23]
范宝磊, 张耀兮, 吴仲珍, 岳霞丽, 索有瑞. 不同品系油菜花粉生活力测定方法比较. 安徽农业科学, 2012, 40: 2596-2597Fan BL, Zhang YX, Wu ZZ, Yue XL, Suo YR. Comparison of different determination methods of the pollen vitality of different lines rape. J Anhui Agric Sci, 2012, 40: 2596-2597 (in Chinese with English abstract)[本文引用:1]
[24]
杜培, 张新友, 李丽娜, 黄冰艳, 易明林, 董文召, 汤丰收. 高质量花生根尖细胞染色体制片方法研究. 河南农业科学, 2013, 42(3): 31-35DuP, Zhang XY, Li LN, Huang BY, Yi ML, Dong WZ, Tang FS. Study on slide preparation methods for high quality chromosomes for root tip cell of Arachis. J Henan Agric Sci, 2013, 42(3): 31-35 (in Chinese with English abstract)[本文引用:1]
[25]
RobledoG, Lavia GI, SeijoG. Species relations among wild Arachis species with the A genome as revealed by FISH mapping of rDNA loci and heterochromatin detection. Theor Appl Genet, 2009, 118: 1295-1307[本文引用:1]
[26]
Stalker HT, Wynne JC. Cytology of interspecific hybrid in section Arachis of peanut. Peanut Sci, 1979, 6: 110-114[本文引用:1]
[27]
StalkerH. T. Utilizing Arachis cardenasii as a source of Cercospora leafspot resistance for peanut improvement. Euphytica, 1984, 33: 529-538[本文引用:1]
[28]
CompanyM, Stalker HT, Wynne JC. Cytology and leafspot resistance in Arachis hypogaea × wild species hybrids. Euphytica, 1982, 31: 885-893[本文引用:1]
[29]
杨小明. 组织培养中秋水仙素诱导葡萄多倍体研究, 甘肃农业大学硕士学位论文, 甘肃兰州, 2003. p 6Yang XM. Studies on Polyploidy Grapevine Induction by Colchicine Treatment Through Tissue Culture. MS Thesis of Gansu Agricultural University, Lanzhou, China, 2003. p 6 (in Chinese with English abstract)[本文引用:1]
[30]
Lavia GI, Ortiz AM, FernándezA. Karyotypic studies in wild germplasm of Arachis (Leguminosae). Genet Resour Crop Evol, 2009, 56: 755-764[本文引用:1]
[31]
Seijo JG, Lavia GI, FernándezA, KrapovickasA, DucasseD, Moscone EA. Physical mapping of 5S and 18S-25S rRNA genes by FISH as evidences that A. duranensis and A. ipaensis are the wild diploid species involved in the origin of A. hypogaea (Leguminosae). Am J Bot, 2004, 91: 1294-1303[本文引用:1]
[32]
杜培, 刘华, 李丽娜, 秦利, 张忠信, 黄冰艳, 董文召, 汤丰收, 亓增军, 张新友. 基于顺序GISH-FISH花生栽培种的染色体分析. 中国农业科学, 2015, 48: 1854-1863DuP, LiuH, Li LN, QinL, Zhang ZX, Huang BY, Dong WZ, Tang FS, Qi ZJ, Zhang XY. Chromosome analysis of peanut (Arachis hypogaea L. ) based on sequential GISH-FISH. Sci Agric Sin, 2015, 48: 1854-1863 (in Chinese with English abstract)[本文引用:1]
[33]
KrapovickasA, Gregory WC. Taxonomia del genero Arachis (Leguminosae). Bonpland ia, 1994, 8: 1-186[本文引用:1]
[34]
Jiang JM, FriebeB, Gill BS. Recent advances in alien gene transfer in wheat. Euphytica, 1994, 73: 199-212[本文引用:1]
 , 杜培
, 杜培





{kind=link}
{kind=link}
{kind=link}
{kind=link}