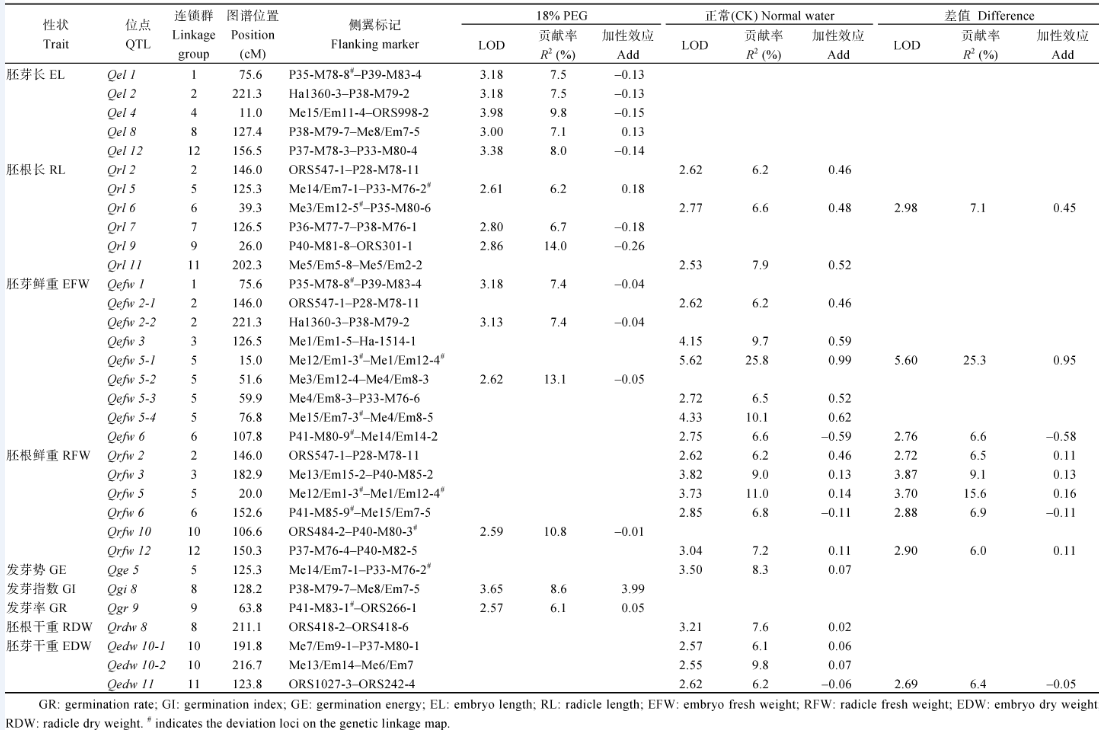
摘要干旱胁迫对向日葵发芽出苗有重要影响。以K55×K58组合衍生的187个F6重组自交系为材料, 利用SSR、SRAP、AFLP标记构建向日葵高密度遗传连锁图谱, 设置正常水分(CK)和模拟干旱(18%聚乙二醇PEG-6000)两种水分条件, 调查9个芽期数量性状, PCR扩增株系, 构建一张包含17个连锁群、1105个标记(368个SSR、368个SRAP和369个AFLP)的高密度遗传连锁图谱。该图谱覆盖基因组长度3846.0 cM, 平均图距3.48 cM, 连锁群长度147.6~295.5 cM, 每个连锁群标记数10~165个。两种条件下检测到33个QTL, 其中干旱条件下检测到发芽指数、发芽率、胚芽长、胚根长、胚芽鲜重和胚根鲜重6个性状的14个QTL, 可解释6.1%~14.0%的表型变异; 正常水分(CK)条件下检测到发芽势、胚根长、胚芽鲜重、胚根鲜重、胚根干重和胚芽干重6个性状的19个QTL, 可解释6.1%~25.8%的表型变异。两种水分条件下检测到 Qefw5-1、Qefw5-2、Qefw5-4、Qrfw5、Qrfw10和 Qrl9共6个QTL的遗传贡献率超过10%, 此外, 还检测到9个影响干旱胁迫与正常水分条件下性状差值的QTL, 可能对抗旱性有直接贡献。这些QTL可为向日葵芽期抗旱分子设计育种研究提供重要参考。
关键词:向日葵; 遗传连锁图谱; 耐旱性; 芽期性状; QTL Construction of High-density Genetic Map and QTL Mapping for Seed Germination Traits in Sunflower under Two Water Conditions LYU Pin1, YU Hai-Feng2, YU Zhi-Xian1, ZHANG Yong-Hu2, ZHANG Yan-Fang2, WANG Ting-Ting1, HOU Jian-Hua1,* 1Inner Mongolia Agricultural University, Huhhot 010010, China
2 Inner Mongolia Academy of Agricultural and Animal Husbandry Sciences, Huhhot 010031, China
Fund:The study was supported by the National Natural Science Foundation of China (31160288, 31540043) AbstractSeed germination and emergence of sunflower are seriously affected by water stress. In this study, SSR, SRAP, and AFLP markers were applied to construct a genetic linkage map by using the F6 population derived from a cross of K55 (drought sensitive) × K58 (drought resistant). For mapping quantitative trait loci (QTLs) for nine traits of seed germination in sunflower, the parents and 187 F6 family lines were used to investigate seed germination traits under normal condition (CK) and 18% polyethylene glycol (PEG-6000) (drought stress). A genetic map consisting of 17 linkage groups was constructed with 1105 loci (368 SSR, 368 SRAP, 369 AFLP) which covers 3846.0 cM and the length of each linkage group varies from 147.6 to 295.5 cM, the number of markers in each linkage group varies from 10 to 165 with an average distance of 3.48 cM. As a result, a total of 33 QTLs were detected. We identified fourteen additive QTLs for germination index (GI), germination rate (GR), embryo length (EL), radicle length (RL), embryo fresh weight (EFW), radicle fresh weight (RFW) under18% PEG condition with explained phenotypic variance ranging from 6.1% to 14.0%. Nineteen additive QTLs were identified for germination energy(GE), radicle length (RL), embryo fresh weight (EFW), radicle fresh weight (RFW), embryo dry weight (EDW), radicle dry weight (RDW) under normal condition with explained phenotypic variance ranging from 6.1% to 25.8%. Each of Qefw5-2, Qefw5-1, Qefw5-4, Qrfw10, Qrfw5, and Qrl9 could explain phenotypic variance over 10%. Nine QTLs affecting trait differences between stress treatment and control were identified, which are considered to directly contribute to drought tolerance. These QTLs identified could provide important reference to molecular breeding for drought-resistance during seed germination in sunflower.
Keyword:Sunflower; Genetic linkage map; Drought tolerance; Seed germination traits; QTL Show Figures Show Figures
刘天鹏, 董孔军, 何继红, 任瑞玉, 张磊, 杨天育. 糜子育成品种芽期抗旱性鉴定与评价研究. 植物遗传资源学报, 2014, 15: 746-752Liu TP, Dong KJ, He JH, Ren RY, ZhangL, Yang TY. Identification and evaluation on the drought resistance of broomcorn millet bred cultivars at germinating stage. J Plant Genet Resour, 2014, 15: 746-752 (in Chinese with English abstract)[本文引用:1]
[2]
Al-Chaarani GR, GentzbittelL, WedzonyM, SarrafiA. Identification of QTLs for germination and seedling development in sunflower (Helianthus annuus L. ). Plant Sci, 2005, 6: 1-7[本文引用:1]
[3]
DavarR, MajdA, DarvishzadehR, SarrafiA. Mapping quantitative trait loci for seedling vigour and development in sunflower (Helianthus annuus L. ) using recombinant inbred line population. Plant Omics J, 2001, 4: 418-427[本文引用:1]
[4]
袁倩倩, 李卓坤, 田纪春, 韩淑晓. 不同水分胁迫下小麦胚芽鞘和胚根长度的QTL分析. 作物学报, 2011, 37: 294-301Yuan QQ, Li ZK, Tian JC, Han SX. QTL Mapping for coleoptile length and radicle length in wheat under different simulated moisture stresses. Acta Agron Sin, 2011, 37: 294-301 (in Chinese with English abstract)[本文引用:1]
[5]
BernardoR. Molecular markers and selection for complex traits in plants: learning from the last 20 years. Crop Sci, 2008, 48: 1649-1664[本文引用:1]
[6]
Rieseberg LH, ChoiH, ChanR. Genomic map of a diploid hybrid species. Heredity, 1993, 70: 285-293[本文引用:1]
Bowers JE, BachlavaE, Brunick RL, Rieseberg LH, Knapp SJ, Burke JM. Development of a 10000 locus genetic map of the sunflower genome based on multiple crosses. Genes Genom Genet, 2012, 2: 721-729[本文引用:1]
[9]
Talukder ZI, GongL, Hulke BS, PegadarajuV, Song QJ, SchultzQ, QiL. A high-density SNP map of sunflower derived from RAD-sequencing facilitating fine-mapping of the rust resistance gene R12. PLoS One, 2014, 7: 1-14[本文引用:1]
[10]
黄先群, GenzbitelleL, FabreF, SaraffiA. SSR分子标记丰富向日葵(Helianthus annuus L. )遗传图谱的研究. 西南农业学报, 2012, 25: 1031-1038Huang XQ, GenzbitelleL, FabreF, SaraffiA. Indication of genetic linkage map for sunflower by SSR markers. Southwest China J Agric Sci, 2012, 25: 1031-1038 (in Chinese with English abstract)[本文引用:1]
[11]
张永虎, 于海峰, 侯建华, 李素萍, 吕品, 于志贤. 利用向日葵重组自交系构建遗传图谱. 遗传, 2014, 36: 1036-1042Zhang YH, Yu HF, Hou JH, Li SP, LyuP, Yu ZX. Construction of a genetic map of sunflower using a population of recombinant inbred lines (RILs), Hereditas, 2014, 36: 1036-1042 (in Chinese with English abstract)[本文引用:3]
[12]
AbdiN, DarvishzadehR, Jafari1 M, Pirzad A, Haddadi P. Genetic analysis and QTL mapping of agro-morphological traits in sunflower (Helianthus annuus L. ) under two contrasting water treatment conditions. Plant Omics J, 2012, 5: 149-158[本文引用:1]
[13]
HaddadiP, Yazdi-samadi B, Naghavi M R, Kalantari A, Maury P, Sarrafi A. QTL analysis of agronomic traits in recombinant inbred lines of sunflower under partial irrigation. Plant Biotechnol Rep, 2011, 5: 135-146[本文引用:1]
[14]
Adiredjo AL, NavaudO, MunosS, Langlade NB, LamazeT, Grieu P. Genetic control of water use efficiency and leaf carbon isotope discrimination in sunflower ( Helianthus annuuL. ) subjected to two drought scenarios. PLoS One , 2014, 101371/journal. pone. 0101218[本文引用:1]
[15]
陈雪, 于海峰, 侯建华, 安玉麟. 向日葵芽期、苗期抗旱性鉴定方法研究. 中国油料作物学报, 2009, 31: 344-348ChenX, Yu HF, Hou JH, An YL. Drought resistance of sunflower in bud and seedling emergences stage. Chin J Oil Crop Sci, 2009, 31: 344-348 (in Chinese with English abstract)[本文引用:1]
[16]
PaniegoN, EchaideM, MuñozM, Fernand ezL, ToralesS, FaccioP, FuxanI, CarreraM, Zand omeniR, Suarez EY, Hopp HE. Microsatellite isolation and characterization in sunflower (Helianthus annuus L. ). Genome, 2002, 45: 34-43[本文引用:1]
[17]
LiG, Quiros CF. Sequence-related amplified polymorphism (SRAP), a new marker system based on a simple PCR reaction: its application to mapping and gene tagging in Brassica. Theor Appl Genet, 2001, 103: 455-461[本文引用:1]
[18]
房冬梅, 吕品, 侯建华. 油葵SSR-PCR反应体系的优化及引物筛选. 中国农学通报, 2015, 31: 205-209Fang DM, LyuP, Hou JH. Optimization of SSR-PCR reaction system and primer screening in oil sunflower. Chin Agric Sci Bull, 2001, 103: 455-461 (in Chinese with English abstract)[本文引用:1]
[19]
于志贤, 侯建华. 油葵SRAP-PCR反应体系的建立与优化. 中国农学通报, 2013, 29: 296-301Yu ZX, Hou JH. Establishment and optimization of SRAP-PCR reaction system in sunflower. Chin Agric Sci Bull, 2013, 29: 296-301 (in Chinese with English abstract)[本文引用:1]
[20]
昝逢刚, 吴才文, 陈学宽, 赵培方, 赵俊, 刘家勇. 118份甘蔗种质资源遗传多样性的AFLP分析. 作物学报, 2014, 40: 1877-1883Zan FG, Wu CW, Chen XK, Zhao PF, ZhaoJ, Liu JY. Genetic diversity of 118 sugarcane germplasm using AFLP markers. Acta Agron Sin, 2014, 40: 1877-1883 (in Chinese with English abstract)[本文引用:1]
[21]
李媛媛, 沈金雄, 王同华, 傅廷栋, 马朝芝. 利用SRAP、SSR和AFLP标记构建甘蓝型油菜遗传连锁图谱. 中国农业科学, 2007, 40: 1118-1126Li YY, Shen JX, Wang TH, Fu TD, Ma CZ. Construction of a linkage map using SRAP, SSR and AFLP markers in Brassica napus L. Sci Agric Sin, 2007, 40: 1118-1126 (in Chinese with English abstract)[本文引用:2]
[22]
吴传书, 王丽侠, 王素华, 陈红霖, 吴健新, 程须珍, 杨晓明. 绿豆高密度分子遗传图谱的构建. 中国农业科学, 2014, 47: 2088-2098Wu CS, Wang LX, Wang SH, Chen HL, Wu JX, Cheng XZ, Yang XM. Construction of a genetic linkage map in mungbean. Sci Agric Sin, 2014, 47: 2088-2098 (in Chinese with English abstract)[本文引用:1]
[23]
王茂芊, 李博, 王华忠. 甜菜遗传连锁图谱初步构建. 作物学报, 2014, 40: 222-230Wang MQ, LiB, Wang HZ. Construction of molecular genetic linkage map of sugarbeet. Acta Agron Sin, 2014, 40: 222-230 (in Chinese with English abstract)[本文引用:1]
[24]
赵秀琴, 朱苓华, 徐建龙, 黎志康. 灌溉与自然降雨条件下水稻高代回交导入系产量QTL的定位. 作物学报, 2007, 33: 1536-1542Zhao XQ, Zhu LH, Xu JL, Li ZK. QTL Mapping of yield under irrigation and rainfed field conditions for advanced backcrossing introgression lines in rice. Acta Agron Sin, 2007, 33: 1536-1542 (in Chinese with English abstract)[本文引用:2]
[25]
彭勃, 王阳, 李永祥, 刘成, 刘志斋, 王迪, 谭巍巍, 张岩, 孙宝成, 石云素, 宋燕春, 王天宇, 黎裕. 不同水分环境下玉米产量构成因子及籽粒相关性状的QTL分析. 作物学报, 2010, 36: 1832-1842PengB, WangY, Li YX, LiuC, Liu ZZ, WangD, Tan WW, ZhangY, Sun BC, Shi YS, Song YC, Wang TY, LiY. QTL Analysis for yield components and kernel-related traits in maize under different water regimes. Acta Agron Sin, 2010, 36: 1832-1842 (in Chinese with English abstract)[本文引用:1]
[26]
谭巍巍, 李永祥, 王阳, 刘成, 刘志斋, 彭勃, 王迪, 张岩, 孙宝成, 石云素, 宋燕春, 杨德光, 王天宇, 黎裕. 在干旱和正常水分条件下玉米穗部性状QTL分析. 作物学报, 2011, 37: 235-248Tan WW, Li YX, WangY, LiuC, Liu ZZ, PengB, WangD, ZhangY, Sun BC, Shi YS, Song YC, Yang DG, Wang TY, LiY. QTL mapping of ear traits of maize under different water regimes. Acta Agron Sin, 2008, 34: 1199-1205 (in Chinese with English abstract)[本文引用:1]
[27]
吴亚辉, 陶星星, 肖武名, 郭涛, 刘永柱, 王慧, 陈志强. 水稻穗部性状的QTL分析. 作物学报, 2014, 40: 214-221Wu YH, Tao XX, Xiao WM, GuoT, Liu YZ, WangH, Chen ZQ. Dissection of QTLs for panicle traits in rice (Oryza sativa). Acta Agron Sin, 2014, 40: 214-221 (in Chinese with English abstract)[本文引用:1]
 , 于海峰
, 于海峰


{kind=link}