关键词:玉米; 穗行数; QTL; 精细定位 Fine Mapping and Genetic Effect Analysis of qKRN5.04, a Major QTL Associated with Kernel Row Number in Maize BAI Na, LI Yong-Xiang*, JIAO Fu-Chao, CHEN Lin, LI Chun-Hui, ZHANG Deng-Feng, SONG Yan-Chun, WANG Tian-Yu, LI Yu, SHI Yun-Su* Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:This study was supported by the Special Fund for Agro-scientific Research in the Public Interest, the Special Fund for Protection and Utilization of Crop Germplasm Resources (201303007, 2015NWB030-04), the National Key Technology Support Program of China (2013BAD01B02-3), the National Natural Science Foundation of China (91335206), and the Agricultural Science and Technology Innovation Program of Chinese Academy of Agricultural Sciences AbstractKernel row number (KRN) is one of the important factors of maize yield. The genetic basis dissection and fine mapping of crucial gene would be greatly beneficial to maize molecular breeding. In present study, series of advanced backcross population and secondary segregation population were developed from a backcross of the derived line of Silunuo (four kernel rows, as the donor parent) and Nong 531 (18-22 kernel rows, as the recurrent parent). The QTL mapping for KRN was conducted by the inclusive complete interval mapping (ICIM) method in multiple environments. And a major KRN QTL, qKRN5.04, was mapped to the interval of 136.3-140.0 Mb on chromosome 5, with the largest phenotypic variation of 21.76% and the effect value of 0.80-1.76 row. Furthermore, according to recombinant analysis of secondary population, qKRN5.04 was fine mapped to the region of about 300 kb, which provided both practical InDel markers for marker-assisted selection and sufficient supports for the map-based cloning and candidate gene mining of the target locus.
Keyword:Maize; Kernel row number (KRN); Quantitative trait locus (QTL); Fine mapping Show Figures Show Figures
表3 玉米穗行数主效位点qKRN5.04的多群体验证和遗传效应分析 Table 3 Effect validation and genetic analysis of multiple populations for major locusqKRN5.04 on kernel row number
环境 Environment
世代 Generation
基因型分析 Genotype analysis
多重比较 Multiple comparison
基因作用 方式 Gene action
农531类型 N531/N531
杂合类型 SLN/N531
四路糯类型 SLN/SLN
农531类型 N531/N531
杂合类型 SLN/N531
四路糯类型 SLN/SLN
2014北京 2014 Beijing
BC3F2
17.24
15.61
14.79
a
b
c
PD
BC4F2
16.91
16.15
15.36
a
b
c
PD
2014石家庄 2014 Shijiazhuang
BC3F2
16.79
15.11
14.53
a
b
c
PD
BC4F2
16.53
15.58
14.70
a
b
c
PD
2014三亚 2014 Sanya
BC3F3
16.56
16.14
15.78
a
a
a
PD
BC4F3
17.66
16.33
15.25
a
ab
b
PD
BC5F2
17.20
16.87
15.90
a
a
b
PD
PD refers to partial dominance effect; different letters in multiple comparison column indicate statistically significant differences among groups (Tukey’ s honestly significant difference (HSD) test, P < 0.05). PD代表部分显性效应; 多重比较(P < 0.05)字母不同代表不同组之间存在显著性差异。
表3 玉米穗行数主效位点qKRN5.04的多群体验证和遗传效应分析 Table 3 Effect validation and genetic analysis of multiple populations for major locusqKRN5.04 on kernel row number
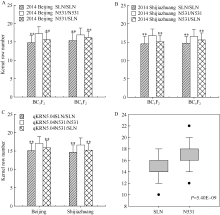
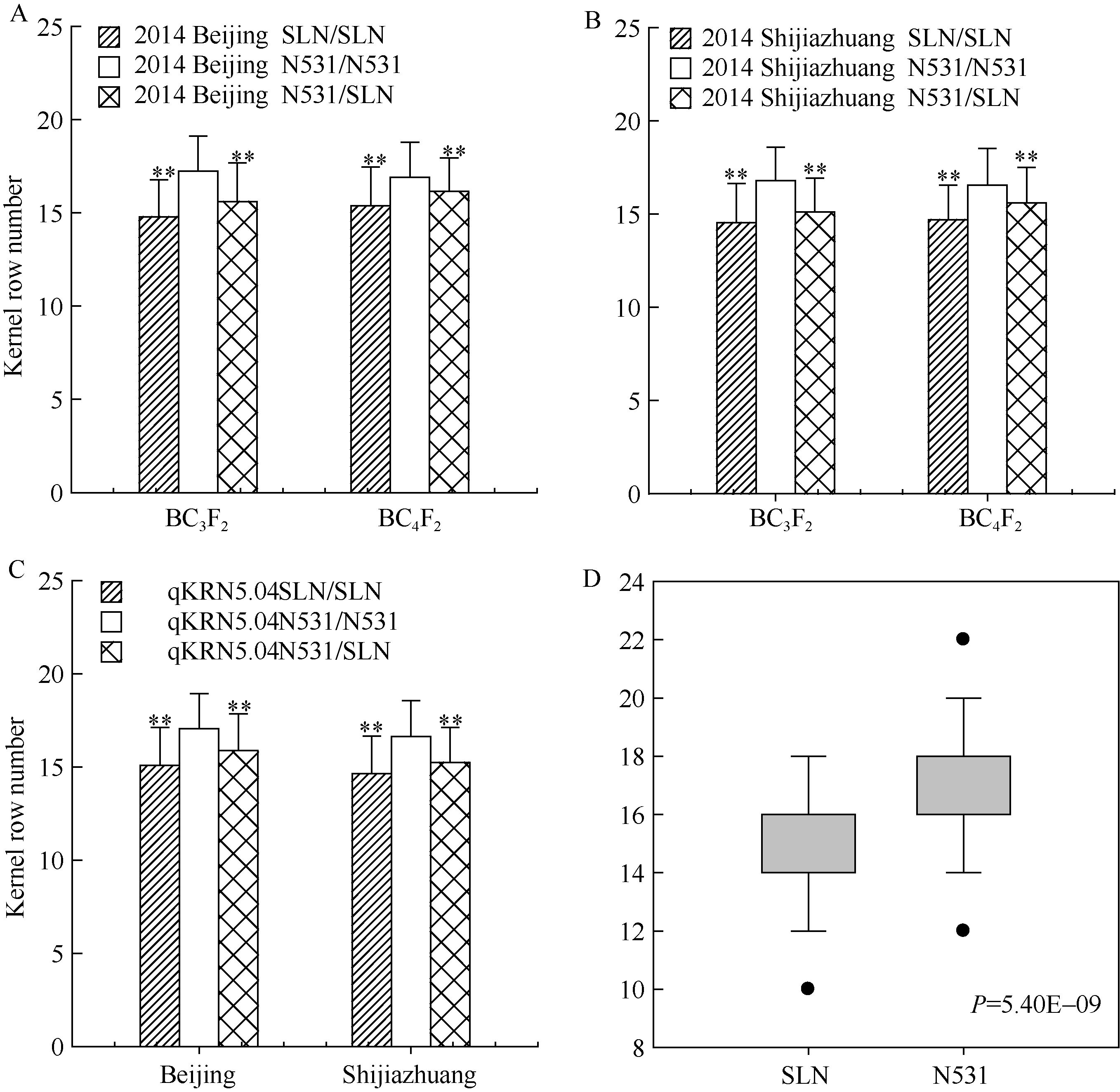
图2 玉米穗行数主效位点qKRN5.04在不同环境条件下的遗传效应比较分析 A, B分别代表2014年北京和河北石家庄环境下的主效位点qKRN5.04在BC3F2和BC4F2群体内的遗传效应比较; C代表2014年北京和河北石家庄2个环境下(群体合并计算) qKRN5.04的遗传效应比较; D代表2014年海南三亚环境下不同基因型BC3F2:3家系间的穗行数差异比较, * * 在代表水平P< 0.01差异极显著; N531、SLN分别是农531和四路糯选系的缩写。Fig. 2 Genetic effect comparisons for kernel row number related major QTL (qKRN5.04) under different environments A, B represent the genetic effects analysis ofqKRN5.04(BC3F2 and BC4F2 population) in 2014 Beijing and 2014 Shijiazhuang, respectively; C represents the genetic effects analysis ofqKRN5.04in 2014 Beijing and 2014 Shijiazhuang (total populations); D represents the KRN comparison of different genotypes of BC3F2:3 family in 2014 Hainan, * * represents significant difference at P< 0.01; N531 and SLN are the abbreviation of Nong 531 and Silunuo, respectively.
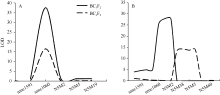
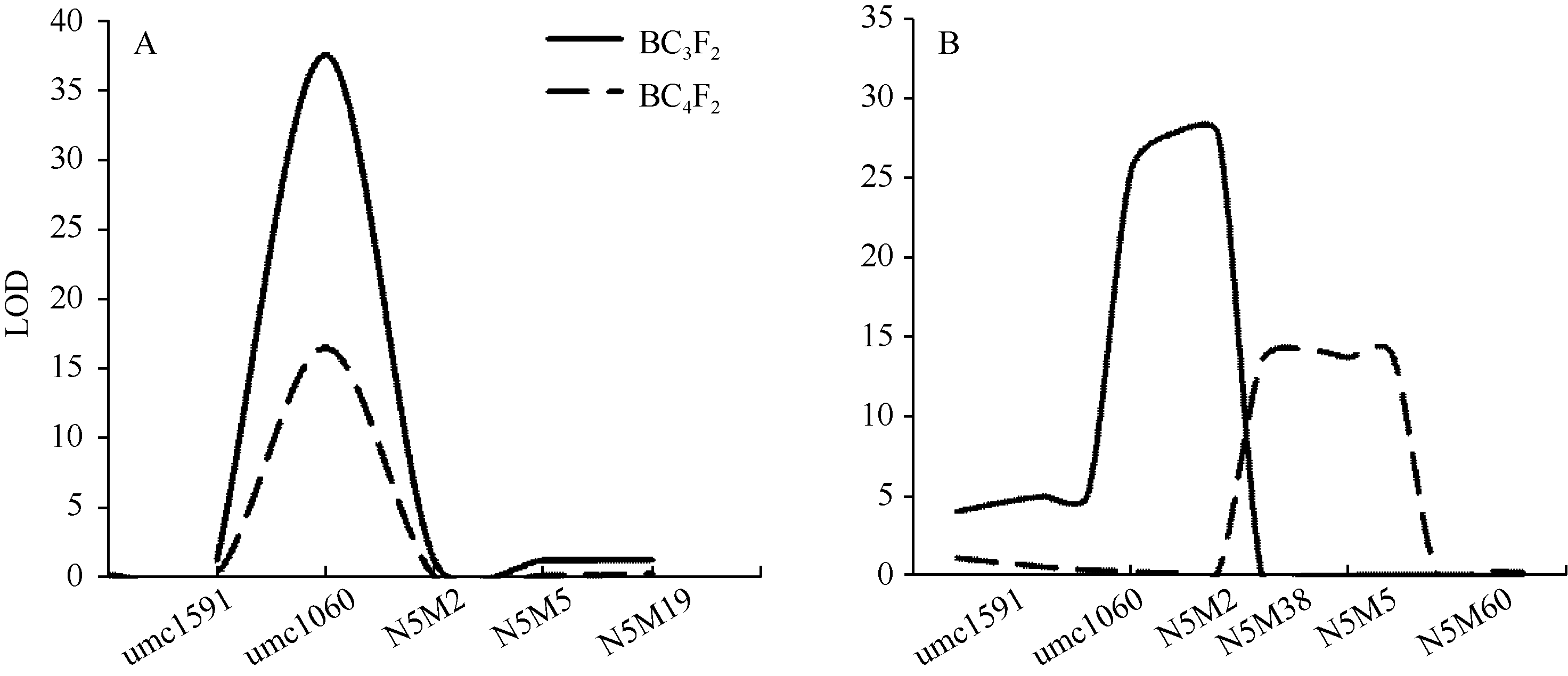
图3 不同环境条件下BC3F2和BC4F2群体的玉米穗行数主效位点qKRN5.04定位 图A和B分别代表qKRN5.04在2014年北京和2014年石家庄的定位结果。Fig. 3 QTL mapping of qKRN5.04using BC3F2 and BC4F2 populations in different environments A and B represent the QTL mapping for qKRN5.04under Beijing and Shijiazhuang in 2014, respectively.
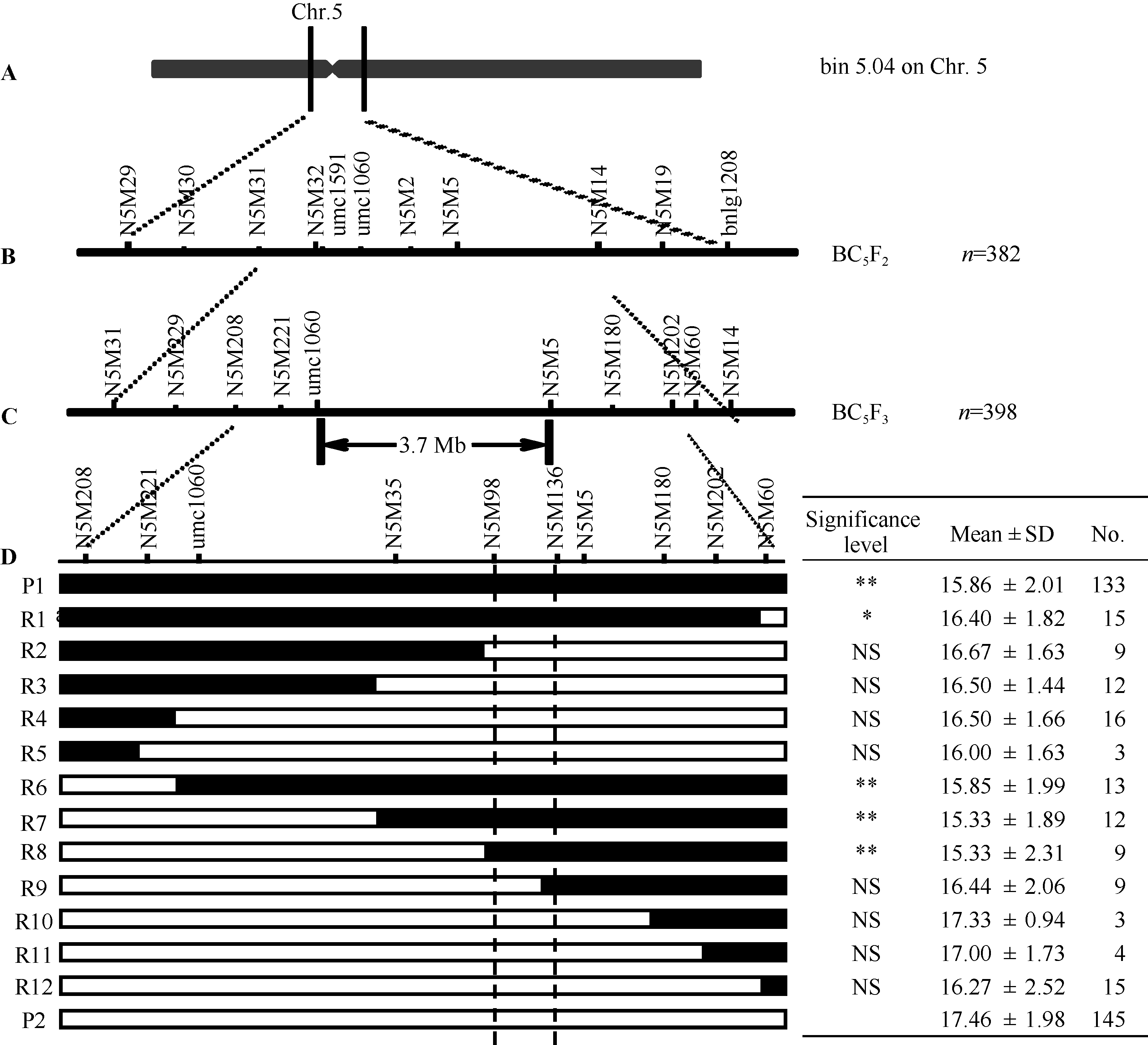
图4 穗行数主效位点qKRN5.04的精细定位 a表示主效位点初定位区段, b和c分别表示BC5F2和BC5F3群体的定位结果, d表示亲本和12种重组类型, 表格表示每种重组类型相应的穗行数和单株个数, 黑色和白色分别代表来自“ 四路糯” (P1)和来自“ 农531” (P2)的染色体片段, * * 表示在0.01水平显著, * 表示在0.05水平显著。Fig. 4 Fine mapping of major QTL qKRN5.04 for kernel row number a shows bin5.04 of Chr.5; b shows qKRN5.04 in BC5F2, and c shows qKRN5.04 in BC5F3, d indicates nine recombinant types (R1-R12) and two parents, and the table shows their corresponding kernel row number, with numbers of individuals for each recombinant on the right. Black and white bars indicate chromosomal segments from SLN (P1) and N531 (P2), respectively. * * and * mean significant difference at the P < 0.01 and 0.05, respectively.
Li JZ, Zhang ZW, Li YL, Wang QL, Zhou YG. QTL consistency and meta-analysis for grain yield components in three generations in maize. Theor Appl Genet, 2011, 122: 771-782[本文引用:1]
[2]
李成璞, 白苇, 翟立红, 陶勇生, 张祖新. 玉米穗行数QTL及其互作分析. 植物遗传资源学报, 2011, 12: 965-970Li CP, BaiW, Zhai LH, Tao YS, Zhang ZX. Identifying and interaction assay of QTL for row number per ear of maize. J Plant Genet Resour, 2011, 12: 965-970 (in Chinese with English abstract)[本文引用:1]
[3]
周强, 王平喜, 程备久, 朱苏文, 谢传晓. 玉米穗行数性状QTL的元分析. 玉米科学, 2014, 22(2): 35-40ZhouQ, Wang PX, Cheng BJ, Zhu SW, Xie CX. Meta-analysis of QTL for ear row number in maize. J Maize Sci, 2014, 22(2): 35-40 (in Chinese with English abstract)[本文引用:2]
[4]
谭巍巍, 李永祥, 王阳, 刘成, 刘志斋, 彭勃, 王迪, 张岩, 孙宝成, 石云素, 宋燕春, 杨德光, 王天宇, 黎裕. 在干旱和正常水分条件下玉米穗部性状QTL分析. 作物学报, 2011, 37: 235-248Tan WW, Li YX, WangY, LiuC, Liu ZZ, PengB, WangD, ZhangY, Sun BC, Shi YS, Song YC, Yang DG, Wang TY, LiY. QTL mapping of ear traits of maize under different water regimes. Acta Agron Sin, 2011, 37: 235-248 (in Chinese with English abstract)[本文引用:1]
[5]
Yan JB, TangH, Huang YQ, Zheng YL, Li JS. Quantitative trait loci mapping and epistatic analysis for grain yield and yield components using molecular markers with an elite maize hybrid. Euphytica, 2006, 149: 121-131[本文引用:2]
[6]
BarazeshS, McSteen P. Barren inflorescence1 functions in organogenesis during vegetative and inflorescence development in maize. Genetics, 2008, 179: 389-401[本文引用:1]
[7]
BarazeshS, NowbakhtC, McSteen P. Sparse inflorescence1, Barren inflorescence1 and Barren stalk1 promote cell elongation in maize inflorescence development. Genetics, 2009, 182: 403-406[本文引用:1]
[8]
Wu XT, SkirpanA, McSteen P. Suppressor of sessile spikelets1 functions in the ramosa pathway controlling meristem determinacy in maize. Plant Physiol, 2009, 149: 205-219[本文引用:1]
[9]
Taguchi-ShiobaraF, YuanZ, HakeS, JacksonD. The fasciated ear2 gene encodes a leucine-rich repeat receptor-like protein that regulates shoot meristem proliferation in maize. Genes Dev, 2001, 15: 2755-2766[本文引用:1]
[10]
VollbrechtE, Springer PS, GohL, Buckler ES, MartienssenR. Architecture of floral branch systems in maize and related grasses. Nature, 2005, 436: 1119-1126[本文引用:1]
[11]
ClermontY, BortiriE. Ramosa2 encodes a lateral organ boundary domain protein that determines the fate of stem cells in branch meristems of maize. Plant Cell, 2006, 18: 574-585[本文引用:1]
[12]
ChuckG, MuszynskiM, KelloggE, HakeS, Schmidt RJ. The control of spikelet meristem identity by the branched silkless1 gene in maize. Science, 2002, 298: 1238-1241[本文引用:1]
[13]
Kaplinsky NJ, FreelingM. Combinatorial control of meristem identity in maize inflorescences. Development, 2003, 130: 1149-1158[本文引用:1]
[14]
BombliesK, Wang RL, Ambrose BA, Schmidt RJ, Meeley RB, DoebleyJ. Duplicate FLORICAULA/LEAFY homologs Zfl1 and Zfl2 control inflorescence architecture and flower patterning in maize. Development, 2003, 130: 2385-2395[本文引用:1]
[15]
周红菊, 穆俊祥, 赵胜杰, 余四斌. 水稻高世代回交导入系耐盐性的遗传研究. 分子植物育种, 2005, 3: 716-720Zhou HJ, Mu JX, Zhao SJ, Yu SB. Genetic analyses of salt tolerance in an advanced backcross population of rice. Mol Plant Breed, 2005, 3: 716-720 (in Chinese with English abstract)[本文引用:1]
[16]
胡利宗, 刘均革, 郭晋杰, 赵永锋, 祝丽英, 宋占权, 陈景堂. 基于玉米BC2F2群体的穗部性状QTL分析. 华北农学报, 2010, 25(4): 73-77Hu LZ, Liu JG, Guo JJ, Zhao YF, Zhu LY, Song ZQ, Chen JT. QTL analysis of ear traits based on BC2F2 population in maize (Zea may L. ). Acta Agr Boreali-Sin, 2010, 25(4): 73-77 (in Chinese with English abstract)[本文引用:1]
[17]
李卫华, 王洪秋, 袁亮, 张向歌, 谢慧玲, 胡彦民, 汤继华. 利用单片段代换系群体定位玉米穗部性状的QTL. 河南农业大学学报, 2013, 47: 143-146Li WH, Wang HQ, YuanL, Zhang XG, Xie HL, Hu YM, Tang JH. Mapping of the QTL for ear related traits using a series of single segment substitution lines in maize. J Henan Agric Univ, 2013, 47: 143-146 (in Chinese with English abstract)[本文引用:1]
[18]
齐欢欢, 段利超, 胡伟, 黄娟, 冯阳, 黄亚群, 祝丽英, 张祖新, 岳兵. 利用导入系群体对玉米产量及产量相关性状进行定位分析. 玉米科学, 2013, 21(4): 24-27Qi HH, Duan LC, HuW, HuangJ, FengY, Huang YQ, Zhu LY, Zhang ZX, YueB. Identification of significant loci for yield and yield-related traits in maize with introgression lines. J Maize Sci, 2013, 21(4): 24-27 (in Chinese with English abstract)[本文引用:1]
[19]
LiF, Jia HT, LiuL, Zhang CX, Liu ZJ, Zhang ZX. Quantitative trait loci mapping for kernel row number using chromosome segment substitution lines in maize. Genet Mol Res, 2014, 13: 1707-1716[本文引用:1]
[20]
Tian BH, Wang JH, Wang GY. Confirmation of a major QTL on chromosome 10 for maize kernel row number in different environments. Plant Breed, 2014, 133: 184-188[本文引用:1]
[21]
LiuL, Du YF, Shen XM, Li MF, SunW, HuangJ, Liu ZL, Tao Y S Zheng Y L, Yan J B, Zhang Z X. KRN4 controls quantitative variation in maize kernel row number. PLoS Genet, 2015, 11(11): e1005670[本文引用:2]
[22]
焦付超, 李永祥, 陈林, 刘志斋, 石云素, 宋燕春, 张登峰, 黎裕, 王天宇. 特异玉米种质四路糯的穗行数遗传解析. 中国农业科学, 2014, 47: 1256-1264Jiao FC, Li YX, ChenL, Liu ZZ, Shi YS, Song YC, Zhang DF, LiY, Wang TY. Genetic dissection for kernel row number in the specific maize germplasm four-rowed waxy corn. Sci Agric Sin, 2014, 47: 1256-1264 (in Chinese with English abstract)[本文引用:2]
[23]
Chen DH, Ronald PC. A rapid DNA min preparation method suitable for AFLP and other PCR applications. Plant Mol Biol Rep, 1999, 17: 53-57[本文引用:1]
[24]
LiH, Ye GJ, Wang JK. A modified algorithm for the improvement of composite interval mapping. Genetics, 2007, 175: 361-374[本文引用:1]
[25]
Stuber CW, Edwards M D. Wendel J F. Molecular marker facilitated investigations of quantitative trait loci in maize: II. Factors influencing yield and its component traits. Crop Sci, 1987, 27: 639-648[本文引用:1]
[26]
谭巍巍, 王阳, 李永祥, 刘成, 刘志斋, 彭勃, 王迪, 张岩, 孙宝成, 石云素, 宋燕春, 杨德光, 王天宇, 黎裕. 不同环境下多个玉米穗部性状的QTL分析. 中国农业科学, 2011, 44: 233-244Tan WW, WangY, Li YX, LiuC, Liu ZZ, PengB, WangD, ZhangY, Sun BC, Shi YS, Song YC, Yang DG, Wang TY, LiY. QTL analysis of ear traits in maize across multiple environments. Sci Agric Sin, 2011, 44: 233-244 (in Chinese with English abstract)[本文引用:1]
[27]
吕学高, 蔡一林, 陈天青, 徐德林, 王伟林, 刘志斋, 王久光. 玉米穗部性状QTL定位. 西南大学学报(自然科学版). 2008, 30(2): 64-70Lyu XG, Cai YL, Chen TQ, Xu DL, Wang WL, Liu ZZ, Wang JG. QTL mapping for ear traits in maize (Zea mays L. ). J Southwest Univ (Nat Sci Edn), 2008, 30(2): 64-70 (in Chinese with English abstract)[本文引用:1]
[28]
Zhuang JY, Lin HX, LuJ, Qian HR, HittalmaniS, HuangN, Zheng KL. Analysis of QTL × environment interaction for yield components and plant height in rice. Theor Appl Genet, 1997, 95: 799-808[本文引用:1]
[29]
FultonT. RFLP mapping of the rice genome. In: Rice Genetics: II. Proceedings of the Second International Rice Genetics Symposium, 14-18 May1990. 2015. pp 435-442[本文引用:1]
[30]
毛传澡, 程式华. 水稻农艺性状QTL定位精确性及其影响因素的分析. 农业生物技术学报, 1999, 7: 386-394Mao CZ, Cheng SH. Analysis of accuracy and influence factor in QTL mapping about agronomic traits in rice (Oryza sativa L. ). J Agric Biol, 1999, 7: 386-394 (in Chinese with English abstract)[本文引用:1]
[31]
Howell PM, Lydiate DJ, Marshall DF. Towards developing intervarietal substitution lines in Brassica napus using marker-assisted selection. Genome, 1996, 39: 348-358[本文引用:1]
[32]
Zhang GD, Wang XP, WangB, Tian YC, LiM, Nie YX, Peng QC, Wang ZL. Fine mapping a major QTL for kernel number per row under different phosphorus regimes in maize (Zea mays L. ). Theor Appl Genet, 2013, 126: 1545-1553[本文引用:1]
[33]
TengF, Zhai LH, Liu RX, BaiW, Wang LQ, Huo DG, Tao YS, Zheng YL, Zhang ZX. ZmGA3ox2, a cand idate gene for a major QTL, qPH3. 1, for plant height in maize. Plant J, 2013, 73: 405-416[本文引用:1]
[34]
Heyndrickx KS, KlaasV. Systematic identification of functional plant modules through the integration of complementary data sources. Plant Physiol, 2012, 159: 884-901[本文引用:1]
[35]
BolducN, YilmazA, Mejia-Guerra M K, Morohashi K, O’Connor D, Grotewold E, Hake S. Unraveling the KNOTTED1 regulatory network in maize meristems. Genes Dev, 2012, 26: 1685-1690[本文引用:2]
 , 李永祥
, 李永祥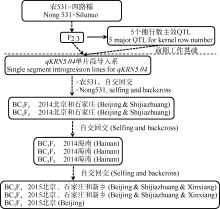


{kind=link}
{kind=link}
{kind=link}
{kind=link}