关键词:水稻; ALK基因; 遗传多样性; 哈尼梯田; 糊化温度 Genetic Diversity of ALK Gene and Its Association with Grain Gelatinization Temperature in Currently Cultivated Rice Landraces from Hani’s Terraced Fields in Yunnan Province LI Chuang1,2,**, LIU Cheng-Chen1,2,**, ZHANG Chang-Quan1, ZHU Ji-Hui1, XU Xiao-Ying2, ZHAO Fu-Wei3, HUANG Shao-Wen4, JIN Yin-Gen2,*, LIU Qiao-Quan1,* 1 Jiangsu Key Laboratory of Crop Genetics and Physiology / Co-Innovation Center for Modern Production Technology of Grain Crops, Yangzhou University, Yangzhou 225009, China
2 College of Biological Science and Technology, Yangzhou University, Yangzhou 225009, China
3 Nanjing Institute of Environmental Sciences of Ministry of Environmental Protection, Nanjing 210042, China
4 Yunnan Honghe University, Mengzi 661100, China
Fund:This study was supported by the National Basic Research Program of China (2013CBA01402), the National Natural Science Foundation of China (31561143008, 31300324), and the Natural Sciences Research Project of Higher Learning Institution in Jiangsu Province (13KJB180028). AbstractYunnan province is one of the centers for genetic diversity of cultivated rice in Asia. A total of 111 rice landraces were collected from Hani’s rice terraces of Yunnan, and their population structure was analyzed by using 51 SSR markers. The 111 rice landraces were divided into japonicagroup (group I) and indicagroup (group II); however, each of the two groups was not completely clustered according to its geographical origin. Most collected landraces, accounting for 83%, belonged to indica group. Gelatinization temperature (GT) is an important parameter affecting rice cooking and eating quality. The GT of rice flour was measured, which was widely different among the collected rice varieties, showing four classes in very low ( <66°C), low (66-70°C), high (70-74°C) and very high ( >74°C) GT. Meanwhile, ALK gene, also known as SSSIIa and controlling GT, was sequenced and compared, showing ten haploid types of the ALKgene among tested samples. Association analysis, revealed that GT is mainly controlled by the combination of three SNPs of ALK gene. Among them, the A-GC or G-TT combination type controls the very low and low GT, while the combination of G-GC controls high and very high GT. SNP3402 type T is associated with high and very high GT. It was also noticed that most of the varieties from indicagroup tended to contain the G-GC mutation and thus showed a relative high GT compared with those from the japonicagroup. These results are meaningful for rice molecular breeding and germplasm resources protection.
Keyword: Oryza sativa L.; ALK gene; Genetic diversity; Hani’; s terrace; Gelatinization temperature Show Figures Show Figures
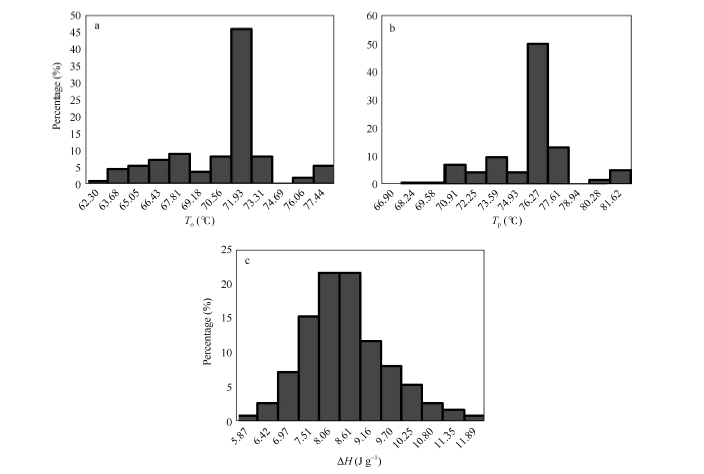
图2 供试样品稻米的糊化温度分布a~c分别为起始糊化温度(To)、最高糊化温度(Tp)和热焓值(Δ H)。Fig. 2 Distribution of gelatinization temperature among tested samplesa-c are onset temperature (To), peak temperature (Tp), and enthalpy of gelatinization (Δ H), respectively.
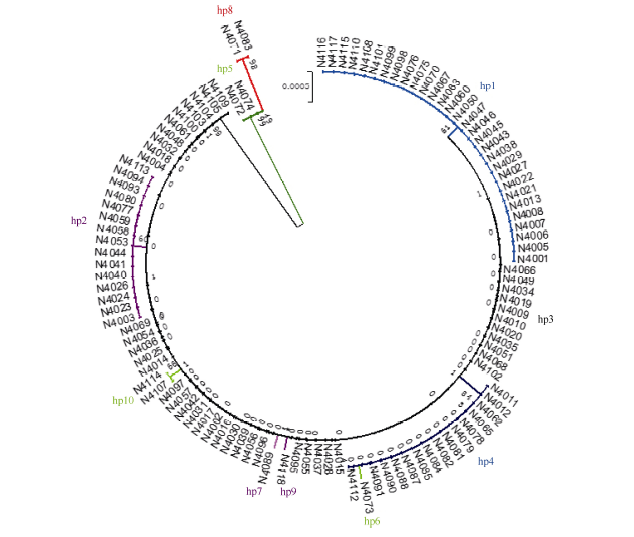
表2ALK基因核苷酸多态性及其与起始糊化温度(To)的关联分析 Table 2 DNA polymorphisms of ALKgene and their association with gelatinization temperature To
区域 Region
位点 Site
Hp1 (30)
Hp2 (15)
Hp3 (42)
Hp4 (15)
Hp5 (2)
Hp6 (1)
Hp7 (1)
Hp8 (2)
Hp9 (1)
Hp10 (2)
Toe
Exon1
185
G
G
G
G
G
G
G
G
A
G
0.019
264
C
C
C
C
C
C
C
G
C
C
0.005
Intron1
412
A
A
A
A
A
A
A
G
A
A
0.005
457
C
C
C
C
C
C
C
T
C
C
0.005
Exon2
516
T
T
T
T
T
T
T
C
T
T
0.005
613
G
G
G
G
G
G
G
G
G
C
0
Intron3
1381
G
G
G
G
G
G
T
G
G
G
0.012
1726
C
C
C
C
A
C
C
A
C
C
0.005
1760
G
G
G
G
C
G
G
C
G
G
0.005
1789
T
T
T
T
C
T
T
C
T
T
0.005
1798
T
T
T
T
C
T
T
C
T
T
0.005
1826
G
G
G
G
A
G
G
A
G
G
0.005
1869
—
—
—
—
T
—
—
T
—
—
0
1915
G
G
G
G
A
G
G
A
G
G
0.005
Intron6
2480
T
T
T
T
A
T
T
A
T
T
0.005
2739
C
C
C
C
A
C
C
A
C
C
0.005
Intron7
3003
G
G
G
G
T
G
G
T
G
G
0.005
2164
A
A
A
A
C
A
A
C
A
A
0.005
3168
C
C
C
C
C
G
C
C
C
C
0
3170
C
C
C
C
T
C
C
T
C
C
0.005
3176
C
C
C
C
A
C
C
A
C
C
0.005
3233
C
C
C
C
G
C
C
G
C
C
0.005
3239
C
C
C
C
A
C
C
A
C
C
0.005
3338
G
G
G
G
A
G
G
A
G
G
0.005
3402
T
C
C
C
C
C
C
C
C
C
0.093* *
3414
G
G
G
G
T
G
G
T
G
G
0.005
3437
—
—
—
—
A
—
—
A
—
—
0
Exon8
3799
A
A
A
A
G
A
A
G
A
A
0.005
3903
T
T
T
T
G
T
T
G
T
T
0.005
4198
G
A
G
G
G
G
G
G
G
G
0.211* * *
4329
G
G
G
T
G
T
G
G
G
G
0.089* *
4330
C
C
C
T
C
T
C
C
C
C
0.089* *
* * Significant at the 0.01 probability level. * * * Significant at the 0.0001 probability level.eCorrelation (R2) betweenALK nucleotide polymorphisms and To. * * 表示在0.01水平差异显著, * * * 表示在0.0001水平差异显著。e核苷酸多态性与To的关联性(R2)。
表2ALK基因核苷酸多态性及其与起始糊化温度(To)的关联分析 Table 2 DNA polymorphisms of ALKgene and their association with gelatinization temperature To
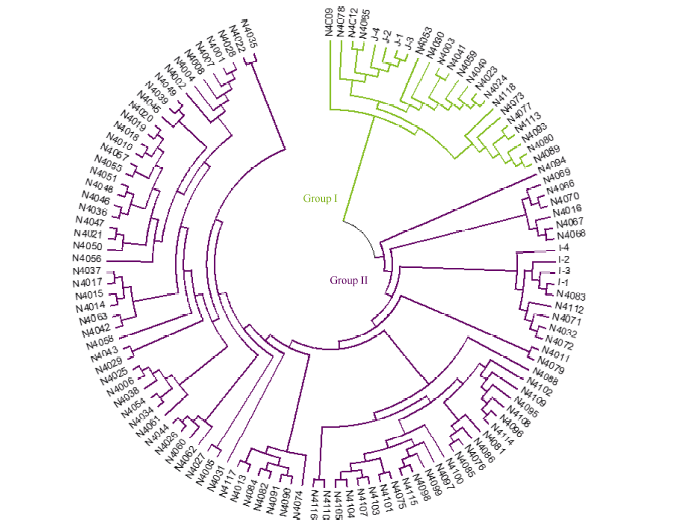
刘承晨, 赵富伟, 吴晓霞, 张昌泉, 朱孔志, 薛达元, 武建勇, 黄绍文, 徐小颖, 金银根, 刘巧泉. 云南哈尼梯田当前栽培水稻遗传多样性及群体结构分析. 中国水稻科学, 2015, 29: 28-34Liu CC, Zhao FW, Wu XX, Zhang CQ, Zhu KZ, Xue DY, Wu JY, Huang SW, Xu XY, Jin YG, Liu QQ. Genetic diversity and population structure analysis of currently cultivated rice land races from Hani’s terraced fields in Yunnan Province. Chin J Rice Sci, 2015, 29: 28-34 (in Chinese with English abstract)[本文引用:3]
[2]
Zeng YW, Shen SQ, Li ZC, Yang ZY, Wang XK, Zhang HL, Wen GS. Ecogeographic and genetic diversity based on morphological characters of indigenous rice ( Oryza sativa L. ) in Yunnan, China. Gen Res Crop Evol, 2003, 50: 567-577[本文引用:1]
[3]
王象坤, 孙传清. 中国栽培稻的起源与演化研究专集. 北京: 中国农业大学出版社, 1996. pp 1-233Wang XK, Sun CQ. Collections of the Origins and Evolution of Cultivated Rice in China. Beijing: China Agricultural University Press, 1996. pp 1-233(in Chinese)[本文引用:1]
[4]
Manish KP, Rani NS, Madhav MS, Sundaram RM, Varaprasad GS, Sivaranjani A K P, Abhishek B, Kumar G R, Kumar A. Different isoforms of starch-synthesizing enzymes controlling amylose and amylopectin content in rice ( Oryza sativaL. ). Biotech Adv, 2012, 30: 1697-1706[本文引用:1]
[5]
董超, 徐福荣, 杨文毅, 汤翠凤, 张恩来, 杨雅云, 阿新祥, 张斐斐, 卢光德, 王艳, 戴陆园. 云南元阳哈尼梯田水稻地方品种月亮谷的遗传变异分析. 中国水稻科学, 2013, 27: 137-144DongC, Xu FR, Yang WY, Tang CF, Zhang EL, Yang YY, A X X, Zhang F F, Lu G D, Wang Y, Dai L Y. Genetic variation analysis of paddy rice land race of Yuelianggu from Yuanyang Hani’s terraced fields in Yunnan province. Chin J Rice Sci, 2013, 27: 137-144 (in Chinese with English abstract)[本文引用:1]
[6]
董树斌, 卢宝荣, 王云月, 杨慧, 涂敏, 李林. 云南水稻传统品种内的遗传多样性及其维持机制初探. 云南农业大学学报, 2010, 25: 1-9Dong SB, Lu BR, Wang YY, YangH, TuM, LiL. Preliminary studies on the within-varietal genetic diversity and its maintenance of traditional rice from Yunnan. J Yunnan Agric Univ, 2010, 25: 1-9 (in Chinese with English abstract)[本文引用:1]
[7]
Gao ZY, Zeng DL, CuiX, Zhou YH, Yan MX, Huang DN, Li JY, QianQ. Map-based cloning of the ALK gene, which controls the gelatinization temperature of rice. Sci China C(Life Sci), 2003, 46: 661-668[本文引用:2]
[8]
Bao JS, CorkeH, SunM. Nucleotide diversity in starch synthase IIa and validation of single nucleotide polymorphisms in relation to starch gelatinization temperature and other physicochemical properties in rice ( Oryza sativaL. ). Theor Appl Genet, 2006, 113: 1113-1171[本文引用:1]
[9]
NakamuraY, Francisco PB, HosakaY, SatoA, SawadaT, KuboA, FujitaN. Essential amino acid of starch synthase IIa differentiate amylopectin structure and starch quality between japonica and indicarice varieties. Plant Mol Biol Rep, 2005, 58: 213-227[本文引用:1]
UmemotoT, YanoM, SatohH, ShomuraA, NakamuraY. Mapping of a gene responsible for the difference in amylopectin structure between japonica-type and indica-type rice varieties. Theor Appl Genet, 2002, 104: 1-8[本文引用:1]
[12]
Li ZY, Li DH, Du XH, WangH, LarroqueO, Jenkins C L D, Jobling S A, Morell M K. The barley amo1 locus is tightly linked to the starch synthase IIIa gene and negatively regulates expression of granule-bound starch synthetic genes. J Exp Bot, 2011, 62: 5217-5231[本文引用:1]
[13]
孙川, 陈刚, 饶玉春, 张光恒, 高振宇, 刘坚, 鞠培娜, 胡江, 郭龙彪, 钱前, 曾大力. 水稻基因组DNA简易制备方法. 中国水稻科学, 2010, 24: 677-680SunC, ChenG, Rao YC, Zhang GH, Gao ZY, LiuJ, Ju PN, HuJ, Guo LB, QianQ, Zeng DL. A simple method for rapid preparation of rice genomic DNA. Chin J Rice Sci, 2010, 24: 677-680 (in Chinese with English abstract)[本文引用:1]
[14]
Zhang CQ, Zhu LJ, ShaoK, Gu MH, Liu QQ. Toward underlying reasons for rice starches having low viscosity and high amylose: physiochemical and structural characteristics. J Sci Food Agric, 2013, 93: 1543-1551[本文引用:1]
[15]
WeiX, Wang RS, Cao LR, Yuan NN, HuangJ, Qiao WG, Zhang WX, Zeng HL, Yang QW. Origin of Oryza sativa in China inferred by nucleotide polymorphisms of organelle DNA. PLoS One, 2012, 7: e49546[本文引用:1]
[16]
YangF, Chen YL, TongC, HuangY, Xu FF, Li KH, CorkeH, SunM, Bao JS. Association mapping of starch physicochemical properties with starch synthesis-related gene markers in nonwaxy rice ( Oryza sativa L. ). Mol Breed, 2014, 34: 1747-1763[本文引用:1]
[17]
Gao ZY, Zeng DL, Cheng FM, Tian ZX, Guo LB, SuY, Yan MX, JiangH, Dong GJ, Huang YC, HanB, Li JY, QianQ. ALK, the key gene for gelatinization temperature, is a modifier gene for gel consistency in rice. J Integr Plant Biol, 2011, 53: 756-765[本文引用:1]
[18]
ZhouY, Zheng HY, Wei GC, ZhouH, Han YN, Bai XF, Xing YZ, Han YP. Nucleotide diversity and molecular evolution of the ALKgene in cultivated rice and its wild relatives. Plant Mol Biol Rep, 2016, DOI: DOI:10.1007/s11105-016-0975-1[本文引用:1]
[19]
Kharabian-MasoulehA, Waters DL, Reinke RF, WardR, Henry RJ. SNP in starch biosynthesis genes associated with nutritional and functional properties of rice. Sci Rep, 2012, 2: 557[本文引用:1]
[20]
Zhang ZJ, LiM, Fang YW, Liu FC, LuY, Meng QC, Peng JC, Yi XH, Gu MH, Yan CJ. Diversification of the Waxy gene is closely related to variations in rice eating and cooking quality. Plant Mol Biol Rep, 2012, 30: 462-469[本文引用:1]
[21]
HoriY, FujimotoR, SatoY, NishioT. A novel wx mutation caused by insertion of a retrotransposon-like sequence in a glutinous cultivar of rice( Oryza sativa). Theor Appl Genet, 2007, 115: 217-224[本文引用:1]
[22]
肖鹏, 邵雅芳, 包劲松. 稻米糊化温度的遗传与分子机理研究进展. 中国农业科技导报, 2010, 12(1): 23-30XiaoP, Shao YF, Bao JS. Research progress on genetics and molecular mechanism of starch gelatinization temperature of rice grain. J Agric Sci Tech, 2010, 12(1): 23-30 (in Chinese with English abstract)[本文引用:1]
[23]
Hoai T TT, MatsusakaH, ToyosawaY, Suu TD, SatohH, KumamaruT. Influence of single-nucleotide polymorphisms in the gene encoding granule-bound starch synthase I on amylose content in Vietnamese rice cultivars. Breed Sci, 2014, 64: 142-148[本文引用:1]
 , 刘巧泉
, 刘巧泉


{kind=link}
{kind=link}
{kind=link}