关键词:作物基因组; 驯化; 育种; 关键基因; 单元型区段 Characteristics of Genes Selected by Domestication and Intensive Breeding in Crop Plants ZHANG Xue-Yong1,*, MA Lin1, ZHENG Jun2 1Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
2Wheat Research Institute, Shanxi Academy of Agricultural Sciences, Linfen 041000, China
Fund:The Principal Investigator was supported by the National Research and Development Program (2016YFD0100300) AbstractCrop genomics made great progress in last 15-20 years. Second generation sequencing technology has dramatically reduced the cost of genome sequencing, brought life science into the times of genomics, and strongly promoted development of system biology, genetics, breeding and genetic resources. Single nucleotide polymorphism (SNP) and haplotype block analysis are currently widely used for exploring animal and plant genetic resources and breeding. Successful isolation of many important genes helps us elucidate the history of crop domestication and breeding, and predict the future of breeding. It has been changing the breeding concepts and strategies. Most crop cultivars used today have experienced two major steps of harsh artificial selection, i.e., domestication and breeding. Despite so many similarities between domestication and breeding, they are different in some aspects. Domestication relatively affects small regions of plant genome. The dramatic reduction of diversity is usually caused by bottleneck effect. Although the breeding history is only about 100 years, it has brought tremendous alteration in most crop genomes. Breeding targets much more genes than domestication. It is very difficult for further selection of alleles at domestication targeted locus, usually only one allele is kept, which referred as fixed. However, it is not in the case of selecting alleles at breeding targeted locus. Few alleles can be present at the same time in cultivar populations and rotate at the same locus in varieties released in different periods or regions. Frequency of favored alleles in new cultivars has been increased dramatically because of positive selection. In addition, strong artificial selection usually reshapes the sub-genomes in polyploid species, which made them quite distinct from donor’s genomes. Therefore, it would be a good strategy to highlight future breeding through elucidating the basic rule of crop genome and gene in reaction to artificial selection at the targeted regions. Here, we briefly review the current major strategies for dissection of genes, haplotype blocks as well as the major genes targeted in crop domestication and breeding selection. We also give a brief introduction on the mission and strategies for “formation and evolution mechanism of funder genotypes and famous cultivars in major crops”, a newly initiated national key research and development program of China.
Keyword:Crop genome; Domestication; Breeding; Targeted genes; Haplotype block Show Figures Show Figures
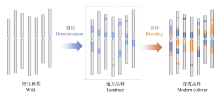
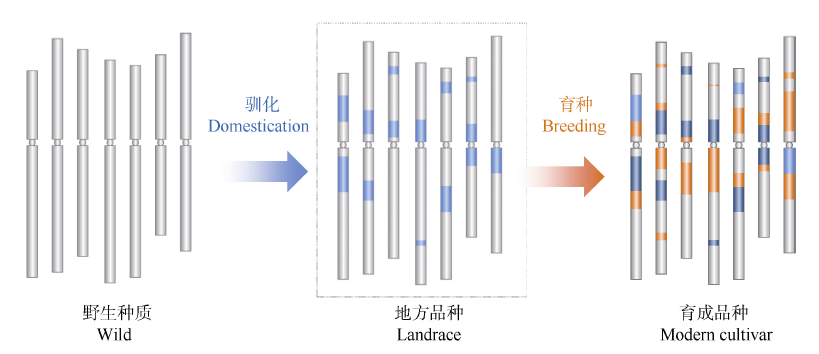
图2 基于选择牵连效应对驯化和育种选择基因组区段的分析(以小麦A基因组为例) 浅蓝色: 驯化选择区段; 深蓝色: 驯化和育种共同选择区段; 橘黄色: 育种选择区段。受选择的区段DNA序列的多样性显著降低。Fig. 2 Identification of haplotype blocks formed in domestication and breeding based on genome resequence and selection sweep analysis (using the A genome of wheat as a model) Bright blue: domestication haplotype blocks; Dark blue:haplotype blocks extensively selected from domestication to breeding; Orange: breeding haplotype blocks. Diversity was dramatically declined at the genomic regions targeted in both domestication and breeding.
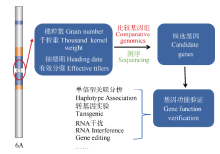
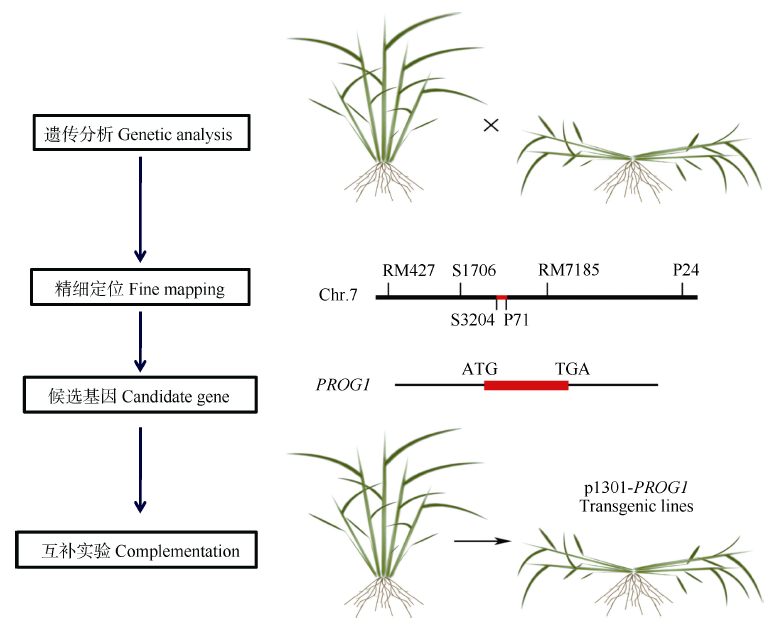
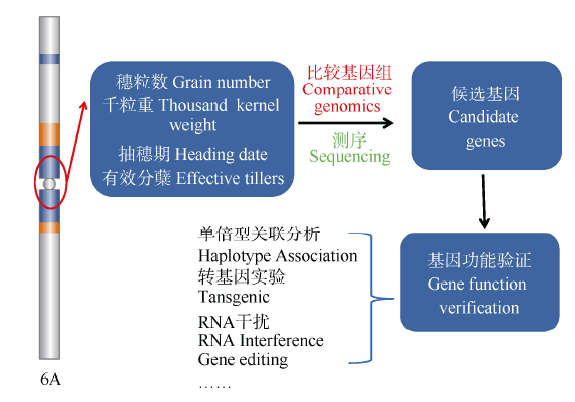
图3 以单元型区段的关联分析为基础, 通过测序和基因组比较分析, 进行关键基因的确认(以小麦6A染色体为例)Fig. 3 Genetic dissection and function verification of the causing genes at the crucial blocks (using wheat 6A as a model)
表1 小麦5A染色体上Vrn1和Vrn2基因的互作与品种冬、春性之关系 Table 1 Gene interaction and genetic decision of winter or spring types of wheat cultivars by two major genes Vrn1and Vrn2 on 5A
基因及基因功能 Gene and gene function
基因互作 Gene interaction
互作效应 Interaction effect
冬春性 Winter/spring type
Vrn1, AP1转录因子, 野生型 Vrn1, AP1 TF, wild type
Vrn1-Vrn2
“ +” 结合, Vrn1转录表达受到抑制 Binding, Vrn1 expression was repressed
冬性Winter type
vrn1, Vrn1突变型, 不转录 vrn1, mutant, no transcription
Vrn1-vrn2
“ -” 无结合, Vrn1正常表达 No binding, Vrn1 expression nomally
春性Spring type
Vrn2, ZCCT转录因子, 野生型 Vrn2, ZCCT TF, wild type
vrn1-Vrn2
“ -” 无结合, vrn1正常表达 No binding, vrn1 expression nomally
“ -” 无结合, vrn1正常表达 No binding, vrn1 expression nomally
春性Spring type
引自Yan L L et al. 2003, 2004[54, 55]. Cited from Yan L L et al. 2003, 2004[54, 55].
表1 小麦5A染色体上Vrn1和Vrn2基因的互作与品种冬、春性之关系 Table 1 Gene interaction and genetic decision of winter or spring types of wheat cultivars by two major genes Vrn1and Vrn2 on 5A
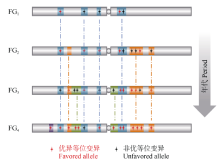
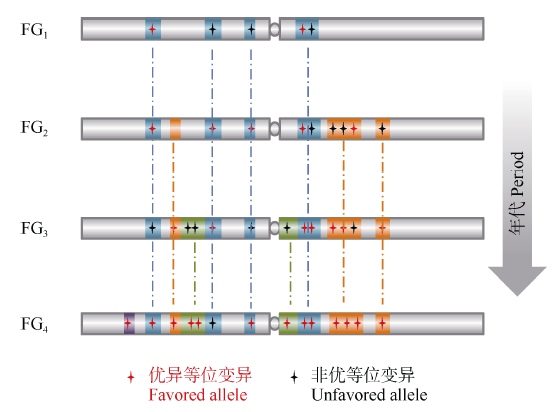
图4 不同时期育种骨干亲本的形成及演替规律解析, 育种的过程就是在驯化基础上对重要基因组区段、基因完善和优化的过程Fig. 4 Strategy for dissection of funder genotypes used at variant periods. Breeding gradually optimizes haplotype blocks and the crucial genes within the blocks
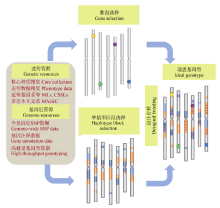
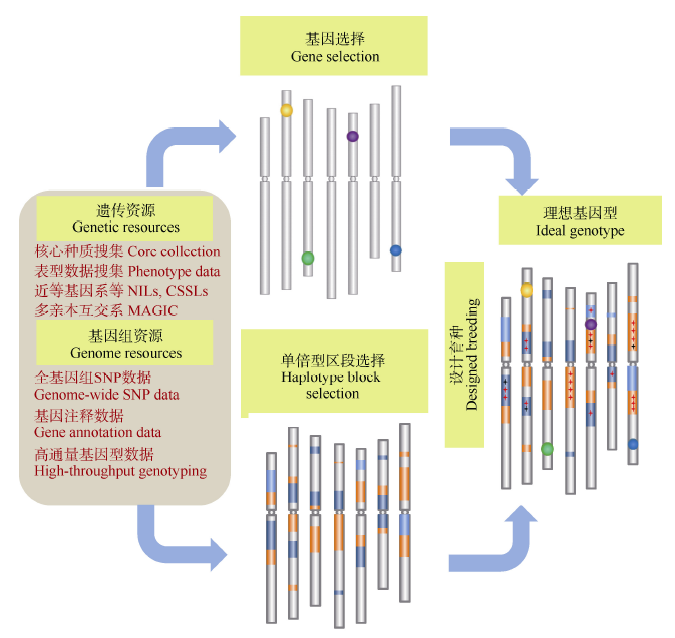
图5 以巢式关联群体(NAM群体)或多亲本符合杂交群体(MAGIC群体)为基础, 通过优良基因与背景的组装, 培育优良品种的基本思路Fig. 5 Strategy for creating ideatype cultivars through integrating important genes and favored haplotype blocks based on the NAM and MAGIC populations
致谢: 感谢万建民院士和徐建龙研究员提出的宝贵意见和建议。因篇幅所限, 只引用了部分同行专家的相关文章, 敬请谅解。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Ross-IbarraJ, Morrell PL, Gaut BS. Plant domestication, a unique opportunity to identify the genetic basis of adaptation. Proc Natl Acad Sci USA, 2007, 104: 8641-8648[本文引用:2]
[3]
Purugganan MD, Fuller DQ. The nature of selection during plant domestication. Nature, 2009, 457: 843-848[本文引用:1]
[4]
IzawaT, KonishiS, ShomuraA, YanoM. DNA changes tell us about rice domestication. Curr Opin Plant Biol, 2009, 12: 185-192[本文引用:1]
[5]
Ge HM, You GX, Wang LF, Hao CY, Dong YS, Li ZS, Zhang XY. Genome selection sweep and association analysis shed light on future breeding by design in wheat. Crop Sci, 2002, 52: 1218-1228[本文引用:3]
[6]
LinT, Zhu GT, Zhang JH, Xu XY, Yu QH, ZhengZ, Zhang ZH, Lun YY, LiS, Wang XX, Huang ZJ, Li JM, Zhang CZ, Wang TT, Zhang YY, Wang AX, Zhang YC, LinK, Li CY, Xiong GS, Xue YB, MazzucatoA, CausseM, Fei ZJ, Giovannoni JJ, Chetelat RT, ZamirD, StadlerT, Li JF, Ye ZB, Du YC, Huang SW. Genomic analyses provide insights into the history of tomato breeding. Nat Genet, 2014, 46: 1220-1226[本文引用:3]
[7]
Zhou DG, ChenW, Lin ZC, Chen HD, Wang CR, LiH, Yu RB, Zhang FY, ZhenG, Yi JL, Li KH, Liu YG, TerzaghiW, Tang XY, HeH, Zhou SC, Deng XW. Pedigree-based analysis of derivation of genome segments of an elite rice reveals key regions during its breeding. Plant Biotechnol J, 2016, 14: 638-648[本文引用:4]
[8]
Doebley JF, Gaut BS, Smith BD. The molecular genetics of crop domestication. Cell, 2006, 127: 1309-1321[本文引用:4]
[9]
Meyer RS, Purugganan MD. Evolution of crop species: genetics of domestication and diversification. Nat Rev Genet, 2013, 14: 840-852[本文引用:3]
[10]
Zhang XY, Tong YP, You GX, Hao CY, Ge HM, Wang LF, LiB, Dong YS, Li ZS. Hitchhiking effect mapping: a new approach for discovering agronomic important genes. Agric Sci China, 2007, 6: 255-264[本文引用:1]
[11]
Olsen KM, Wendel JF. A bountiful harvest: Genomic insights into crop domestication phenotypes. Annu Rev Plant Biol, 2013, 64(4): 47-70[本文引用:1]
[12]
JinJ, HuangW, Gao JP, YangJ, ShiM, Zhu MZ, LuoD, Lin HX. Genetic control of rice plant architecture under domestication. Nat Genet, 2008, 40: 1365-1369[本文引用:2]
[13]
Tan LB, Li XR, Liu FX, Sun XY, Li CG, Zhu ZF, Fu YC, Cai HW, Wang XK, Xie DX, Sun CQ. Control of a key transition from prostrate to erect growth in rice domestication. Nat Genet, 2008, 40: 1360-1364[本文引用:2]
[14]
Xie WB, Wang GW, YuanM, YaoW, LyuK, ZhaoH, YangM, Li PB, ZhangX, YuanJ, Wang QX, LiuF, Dong HX, ZhangLJ, Li XL, Meng XZ, ZhangW, Xiong LZ, He YQ, Wang SP, Yu SB, Xu CG, LuoJ, Li XH, Xiao JH, Lian XM, Zhang QF. Breeding signatures of rice improvement revealed by a genomic variation map from a large germplasm collection. Proc Natl Acad Sci USA, 2015, 112: 5411-5419[本文引用:2]
[15]
HouJ, Jiang QY, Hao CY, Wang YQ, Zhang HN, Zhang XY. Global selection on sucrose synthase haplotypes during a century of wheat breeding. Plant Physiol, 2014, 164: 1918-1929[本文引用:6]
[16]
ShangY, Ma YS, ZhouY, Zhang HM, Duan LX, Chen HM, Zeng JG, ZhouQ, Wang SH, Gu WJ, LiuM, Ren JW, Gu XF, Zhang SP, WangY, YasukawaK, Bouwmeester HJ, Qi XQ, Zhang ZH, Lucas WJ, Huang SW. Biosynthesis, regulation, and domestication of bitterness in cucumber. Science, 2014, 346: 1084-1088[本文引用:2]
[17]
KonishiS, IzawaT, Lin SY, EbanaK, FukutaY, SasakiT, YanoM. An SNP caused loss of seed shattering during rice domestication. Science, 2006, 312: 1392-1396[本文引用:1]
[18]
Li CB, Zhou AL, SangT. Rice domestication by reducing shattering. Science, 2006, 311: 1936-1939[本文引用:1]
[19]
Lin ZW, Li XR, Shannon LM, Yeh CT, Wang ML, Bai GH, PengZ, Li JR, Trick HN, Clemente TE, DoebleyJ, Schnable PS, Tuinstra MR, Tesso TT, WhiteF, Yu JM. Parallel domestication of the Shattering1 genes in cereals. Nat Genet, 2012, 44: 720-754[本文引用:1]
[20]
Simons KJ, Fellers JP, Trick HN, Zhang ZC, Tai YS, Gill BS, Faris JD. Molecular characterization of the major wheat domestication gene Q. Genetics, 2006, 172: 547-555[本文引用:1]
StuderA, ZhaoQ, Ross-IbarraJ, DoebleyJ. Identification of a functional transposon insertion in the maize domestication gene tb1. Nat Genet, 2011, 43: 1160-1164[本文引用:1]
[23]
Zhou LL, Zhang JY, Yan JB, Song RT. Two transposable element insertions are causative mutations for the major domestication gene teosinte branched 1 in modern maize. Cell Res, 2011, 21: 1267-1270[本文引用:1]
[24]
Tian ZX, Wang XB, LeeR, Li YH, Specht JE, Nelson RL, McClean P E, Qiu L J, Ma J X. Artificial selection for determinate growth habit in soybean. Proc Natl Acad Sci USA, 2010, 107: 8563-8568[本文引用:1]
[25]
Ping JQ, Liu YF, Sun LJ, Zhao MX, Li YH, She MY, SuiY, LinF, Liu XD, Tang ZX, NguyenH, Tian ZX, Qiu LJ, Nelson RL, Clemente TE, Specht JE, Ma JX. Dt2 is a gain-of-function MADS-domain factor gene that specifies semideterminacy in soybean. Plant Cell, 2014, 26: 2831-2842[本文引用:1]
[26]
Zuo JR, Li JY. Molecular genetic dissection of quantitative trait loci regulating rice grain size. Annu Rev Genet, 2014, 48: 99-118[本文引用:3]
[27]
MaL, LiT, Hao CY, Wang YQ, Chen XH, Zhang XY. TaGS5-3A, a grain size gene selected during wheat improvement for larger kernel and yield. Plant Biotechnol J, 2016, 14: 1269-1280[本文引用:2]
[28]
CongB, Barrero LS, Tanksley SD. Regulatory change in YABBY-like transcription factor led to evolution of extreme fruit size during tomato domestication. Nat Genet, 2008, 40: 800-804[本文引用:1]
[29]
Whitt SR, Wilson LM, Tenaillon MI, Gaut BS, Buckler ES. Genetic diversity and selection in the maize starch pathway. Proc Natl Acad Sci USA, 2002, 99: 12959-12962[本文引用:2]
[30]
Ding ZH, Wang CR, ChenS, Yu SB. Diversity and selective sweep in the OsAMT1;1 genomic region of rice. BMC Evol Biol, 2011, 11: 1-10[本文引用:1]
[31]
MaY, Dai XY, Xu YY, LuoW, Zheng XM, Zeng DL, Pan YJ, Lin XL, Liu HH, Zhang DJ, XiaoJ, Guo XY, Xu SJ, Niu YD, Jin JB, ZhangH, XuX, Li LG, WangW, QianQ, GeS, ChongK. COLD1 confers chilling tolerance in rice. Cell, 2015, 160: 1209-1221[本文引用:1]
[32]
Tanksley SD, McCouch S R. Seed banks and molecular maps: unlocking genetic potential from the wild. Science, 1997, 277: 1063-1066[本文引用:1]
[33]
WangE, WangJ, Zhu XD, HaoW, Wang LY, LiQ, Zhang LX, HeW, Lu BR, Lin HX, MaH, Zhang GQ, He ZH. Control of rice grain-filling and yield by a gene with a potential signature of domestication. Nat Genet, 2008, 40: 1370-1374[本文引用:2]
LiuY, Xu JX, Ding YF, Wang QS, Li GH, Wang SH. Auxin inhibits the outgrowth of tiller buds in rice (Oryza sativa L. ) by downregulating OsIPT expression and cytokinin biosynthesis in nodes. Aust J Crop Sci, 2011, 5: 169-174[本文引用:1]
[36]
ZhouF, Lin QB, Zhu LH, Ren YL, Zhou KN, ShabekN, Wu FQ, Mao HB, DongW, GanL, Ma WW, GaoH, ChenJ, YangC, WangD, Tan JJ, ZhangX, Guo XP, Wang JL, JiangL, LiuX, Chen WQ, Chu JF, Yan CY, UenoK, ItoS, AsamiT, Cheng ZJ, WangJ, Lei CL, Zhai HQ, Wu CY, Wang HY, ZhengN, Wan JM. D14-SCFD3-dependent degradation of D53 regulates strigolactone signalling. Nature, 2013, 504: 406-410[本文引用:1]
[37]
JiangL, LiuX, Xiong GS, Liu HH, Chen FL, WangL, Meng XB, Liu GF, YuH, Yuan YD, YiW, Zhao LH, Ma HL, He YZ, Wu ZS, MelcherK, QianQ, Xu HE, Wang YH, Li JY. DWARF53 acts as a repressor of strigolactone signalling in rice. Nature, 2013, 504: 401-405[本文引用:1]
[38]
Li XY, QianQ, Fu ZM, Wang YH, Xiong GS, Zeng DL, Wang XQ, Liu XF, TengS, HiroshiF, YuanM, LuoD, HanB, Li JY. Control of tillering in rice. Nature, 2003, 422: 618-621[本文引用:1]
[39]
Wang WF, LiG, ZhaoJ, Chu HW, Lin WH, Zhang DB, Wang ZY, Liang WQ. DWARF TILLER1, a WUSCHEL-related homeobox transcription factor, is required for tiller growth in rice. PLoS Genet, 2014, 10: e1004154[本文引用:1]
[40]
Xue WY, Xing YZ, Weng XY, ZhaoY, Tang WJ, WangL, Zhou HJ, Yu SB, Xu CG, Li XH, Zhang QF. Natural variation in Ghd7 is an important regulator of heading date and yield potential in rice. Nat Genet, 2008, 40: 761-767[本文引用:2]
[41]
AshikariM, SakakibaraH, Lin SY, YamamotoT, TakashiT, NishimuraA, Angeles ER, QianQ, KitanoH, MatsuokaM. Cytokinin oxidase regulates rice grain production. Science, 2005, 309: 741-745[本文引用:1]
[42]
Li SY, Zhao BR, Yuan DY, Duan MJ, QianQ, TangL, WangB, Liu XQ, ZhangJ, WangJ, Sun JQ, LiuZ, Feng YQ, Yuan LP, Li CY. Rice zinc finger protein DST enhances grain production through controlling Gn1a/OsCKX2 expression. Proc Natl Acad Sci USA, 2013, 110: 3167-3172[本文引用:1]
[43]
LiuL, Du YF, Shen XM, Li MF, SunW, HuangJ, Liu ZJ, Tao YS, Zheng YL, Yan JB, Zhang ZX. KRN4 controls quantitative variation in maize kernel row number. PLoS Genet, 2015, 11: e1005670[本文引用:1]
[44]
ZhengJ, LiuH, Wang YQ, Wang LF, Chang XP, Jing RL, Hao CY, Zhang XY. TEF-7A, a transcript elongation factor gene, influences yield-related traits in bread wheat (Triticum aestivum L. ). J Exp Bot, 2014, 65: 5351-5365[本文引用:1]
[45]
LiuJ, HuaW, Hu ZY, Yang HL, ZhangL, Li RJ, Deng LB, Sun XC, Wang XF, Wang HZ. Natural variation in ARF18 gene simultaneously affects seed weight and silique length in polyploid rapeseed. Proc Natl Acad Sci USA, 2015, 112: 5123-5132[本文引用:1]
[46]
ChalhoubB, DenoeudF, Liu SY, Parkin I A P, Tang H B, Wang X Y, Chiquet J, Belcram H, Tong C B, Samans B, Correa M, Da Silva C, Just J, Falentin C, Koh C S, Le Clainche I, Bernard M, Bento P, Noel B, Labadie K, Alberti A, Charles M, Arnaud D, Guo H, Daviaud C, Alamery S, Jabbari K, Zhao M X, Edger P P, Chelaifa H, Tack D, Lassalle G, Mestiri I, Schnel N, Le Paslier M C, Fan G Y, Renault V, Bayer P E, Golicz A A, Manoli S, Lee T H, Thi V H D, Chalabi S, Hu Q, Fan C C, Tollenaere R, Lu Y H, Battail C, Shen J X, Sidebottom C H D, Wang X F, Canaguier A, Chauveau A, Berard A, Deniot G, Guan M, Liu Z S, Sun F M, Lim Y P, Lyons E, Town C D, Bancroft I, Wang X W, Meng J L, Ma J X, Pires J C, King G J, Brunel D, Delourme R, Renard M, Aury J M, Adams K L, Batley J, Snowdon R J, Tost J, Edwards D, Zhou Y M, Hua W, Sharpe A G, Paterson A H, Guan C Y, Wincker P. Early allopolyploid evolution in the post-neolithic Brassica napus oilseed genome. Science, 2014, 345: 950-953[本文引用:2]
[47]
Peng JR, Richards DE, Hartley NM, Murphy GP, Devos KM, Flintham JE, BealesJ, Fish LJ, Worland AJ, PelicaF, SudhakarD, ChristouP, Snape JW, Gale MD, Harberd NP. ‘Green revolution’ genes encode mutant gibberellin response modulators. Nature, 1999, 400: 256-261[本文引用:1]
[48]
Fu XD, Richards DE, Ait-AliT, Hynes LW, OughamH, Peng JR, Harberd NP. Gibberellin-mediated proteasome-dependent degradation of the barley DELLA protein SLN1 repressor. Plant Cell, 2002, 14: 3191-3200[本文引用:1]
[49]
WuJ, Kong XY, Wan JM, Liu XY, ZhangX, Guo XP, Zhou RH, Zhao GY, Jing RL, Fu XD, Jia JZ. Dominant and pleiotropic effects of a GAI gene in wheat results from a lack of interaction between DELLA and GID1. Plant Physiol, 2011, 157: 2120-2130[本文引用:1]
[50]
杨松杰, 张晓科, 何中虎, 夏先春, 周阳. 用STS标记检测矮秆基因Rht-B1b和Rht-D1b在中国小麦中的分布. 中国农业科学, 2006, 39: 1680-1688Yang SJ, Zhang XK, He ZH, Xia XC, ZhouY. Distrubition of dwarfing genes Rht-B1b and Rht-D1b in Chinese bread wheats detected by STS marker. Sci Agric Sin, 2006, 39: 1680-1688 (in Chinese with English abstract)[本文引用:1]
[51]
AsanoK, TakashiT, MiuraK, QianQ, KitanoH, MatsuokaM, AshikariM. Genetic and molecular analysis of utility of sd1 alleles in rice breeding. Breed Sci, 2007, 57: 53-58[本文引用:1]
[52]
Wu WX, Zheng XM, Lu GW, Zhong ZZ, GaoH, Chen LP, Wu CY, Wang HJ, WangQ, Zhou KN, Wang JL, Wu FQ, ZhangX, Guo XP, Cheng ZJ, Lei CL, Lin QB, JiangL, Wang HY, GeS, Wan JM. Association of functional nucleotide polymorphisms at DTH2 with the northward expansion of rice cultivation in Asia. Proc Natl Acad Sci USA, 2013, 110: 2775-2780[本文引用:1]
[53]
YangQ, LiZ, Li WQ, Ku LX, WangC, Ye JR, LiK, YangN, Li YP, ZhongT, Li JS, Chen YH, Yan JB, Yang XH, Xu ML. CACTA-like transposable element in ZmCCT attenuated photoperiod sensitivity and accelerated the postdomestication spread of maize. Proc Natl Acad Sci USA, 2003, 110: 16969-16974[本文引用:1]
[54]
Yan LL, LoukoianovA, TranquilliG, HelgueraM, FahimaT, DubcovskyJ. Positional cloning of the wheat vernalization gene VRN1. Proc Natl Acad Sci USA, 2003, 100: 6263-6268[本文引用:1]
[55]
Yan LL, LoukoianovA, BlechlA, TranquilliG, RamakrishnaW, SanMiguel P, Bennetzen J L, Echenique V, Dubcovsky J. The wheat VRN2 gene is a flowering repressor down-regulated by vernalization. Science, 2004, 303: 1640-1644[本文引用:1]
[56]
TrevaskisB, Hemming MN, Peacock WJ, Dennis ES. HvVRN2 responds to daylength, whereas HvVRN1 is regulated by vernalization and developmental status. Plant Physiol, 2006, 140: 1397-1405[本文引用:1]
[57]
Guo ZA, Song YX, Zhou RH, Ren ZL, Jia JZ. Discovery, evaluation and distribution of haplotypes of the wheat Ppd-D1 gene. New Phytol, 2010, 185: 841-851[本文引用:1]
[58]
TurnerA, BealesJ, FaureS, Dunford RP, Laurie DA. The pseudo-response regulator Ppd-H1 provides adaptation to photoperiod in barley. Science, 2005, 310: 1031-1034[本文引用:1]
[59]
Staskawicz BJ, Ausubel FM, Baker BJ, Ellis JG, Jones J D G. Molecular-genetics of plant-disease resistance. Science, 1995, 268: 661-667[本文引用:1]
[60]
Song WY, Pi LY, Wang GL, GardnerJ, HolstenT, Ronald PC. Evolution of the rice Xa21 disease resistance gene family. Plant Cell, 1997, 9: 1279-1287[本文引用:1]
[61]
Liu YQ, WuH, ChenH, Liu YL, HeJ, Kang HY, Sun ZG, PanG, WangQ, Hu JL, ZhouF, Zhou KN, Zheng XM, Ren YL, Chen LM, Wang YH, Zhao ZG, Lin QB, Wu FQ, ZhangX, Guo XP, Cheng XI, JiangL, Wu CY, Wang HY, Wan JM. A gene cluster encoding lectin receptor kinases confers broad- spectrum and durable insect resistance in rice. Nat Biotechnol, 2015, 33: 301-305[本文引用:1]
[62]
Cook DE, Lee TG, Guo XL, MelitoS, WangK, Bayless AM, Wang JP, Hughes TJ, Willis DK, Clemente TE, Diers BW, Jiang JM, Hudson ME, Bent AF. Copy number variation of multiple genes at Rhg1 mediates nematode resistance in soybean. Science, 2012, 338: 1206-1209[本文引用:1]
[63]
Krattinger SG, Lagudah ES, WickerT, Risk JM, Ashton AR, Selter LL, MatsumotoT, KellerB. Lr34 multi-pathogen resistance ABC transporter: molecular analysis of homoeologous and orthologous genes in hexaploid wheat and other grass species. Plant J, 2011, 65: 392-403[本文引用:1]
[64]
Moore JW, Herrera-FoesselS, Lan CX, SchnippenkoetterW, AyliffeM, Huerta-EspinoJ, LillemoM, ViccarsL, MilneR, PeriyannanS, Kong XY, SpielmeyerW, TalbotM, BarianaH, Patrick JW, DoddsP, SinghR, LagudahE. A recently evolved hexose transporter variant confers resistance to multiple pathogens in wheat. Nat Genet, 2015, 47: 1494-1498[本文引用:1]
[65]
ChenM, Xu ZS, Xia LQ, Li LC, Cheng XG, Dong JH, Wang QY, Ma YZ. Cold-induced modulation and functional analyses of the DRE-binding transcription factor gene, GmDREB3, in soybean (Glycine max L. ). J Exp Bot, 2009, 60: 121-135[本文引用:1]
[66]
James RA, Davenport RJ, MunnsR. Physiological characterization of two genes for Na+ exclusion in durum wheat, Nax1 and Nax2. Plant Physiol, 2006, 142: 1537-1547[本文引用:1]
[67]
MunnsR, James RA, XuB, AthmanA, Conn SJ, JordansC, Byrt CS, Hare RA, Tyerman SD, TesterM, PlettD, GillihamM. Wheat grain yield on saline soils is improved by an ancestral Na+ transporter gene. Nat Biotechnol, 2012, 30: 360-373[本文引用:2]
[68]
Jia JZ, Zhao SC, Kong XY, Li YR, Zhao GY, He WM, AppelsR, PfeiferM, TaoY, Zhang XY, Jing RL, ZhangC, Ma YZ, Gao LF, GaoC, SpannaglM, Mayer K F X, Li D, Pan S K, Zheng F Y, Hu Q, Xia X C, Li J W, Liang Q S, Chen J, Wicker T, Gou C Y, Kuang H H, He G Y, Luo Y D, Keller B, Xia Q J, Lu P, Wang J Y, Zou H F, Zhang R Z, Xu J Y, Gao J L, Middleton C, Quan Z W, Liu G M, Wang J, Yang H M, Liu X, He Z H, Mao L, Wang J. Aegilops tauschii draft genome sequence reveals a gene repertoire for wheat adaptation. Nature, 2013, 496: 91-95[本文引用:1]
[69]
Yang CW, ZhaoL, Zhang HK, Yang ZZ, WangH, Wen SS, Zhang CY, Rustgi S, von Wettstein D, Liu B. Evolution of physiological responses to salt stress in hexaploid wheat. Proc Natl Acad Sci USA, 2014, 111: 11882-11887[本文引用:1]
[70]
Mickelbart MV, Hasegawa PM, Bailey-SerresJ. Genetic mechanisms of abiotic stress tolerance that translate to crop yield stability. Nat Rev Genet, 2015, 16: 237-251[本文引用:1]
[71]
Wang LF, Ge HM, Hao CY, Dong YS, Zhang XY. Identifying loci influencing 1000-kernel weight in wheat by microsatellite screening for evidence of selection during breeding. PLoS One, 2012, 7: e29432[本文引用:2]
[72]
Zhang DL, Hao CY, Wang LF, Zhang XY. Identifying loci influencing grain number by microsatellite screening in bread wheat (Triticum aestivum L. ). Planta, 2012, 236: 1507-1517[本文引用:2]
[73]
Zhang HK, ZhuB, QiB, Gou XW, Dong YZ, Xu CM, Zhang BJ, HuangW, LiuC, Wang XT, Yang CW, ZhouH, KashkushK, FeldmanM, Wendel JF, LiuB. Evolution of the BBAA component of bread wheat during its history at the allohexaploid level. Plant Cell, 2014, 26: 2761-2776[本文引用:1]
[74]
XuK, XuX, FukaoT, CanlasP, Maghirang-RodriguezR, HeuerS, Ismail AM, Bailey-SerresJ, Ronald PC, Mackill DJ. Sub1A is an ethylene-response-factor-like gene that confers submergence tolerance to rice. Nature, 2006, 442: 705-708[本文引用:2]
[75]
Peleman JD, van der Voort J R. Breeding by design. Trends Plant Sci, 2003, 8: 330-334[本文引用:1]
 , 马琳
, 马琳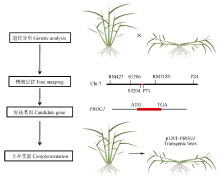

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}