关键词:糜子; 荧光SSR; 遗传多样性; 聚类分析; 遗传结构 Analysis of Genetic Diversity in Common Millet ( Panicum miliaceum) Using Fluorescent SSR in China WANG Rui-Yun1,2,*,**, JI Xu1,**, LU Ping3, LIU Min-Xuan3, XU Yue4, WANG Lun2, WANG Hai-Gang2, QIAO Zhi-Jun2,* 1College of Agriculture, Shanxi Agricultural University, Taigu 030801, China
2 Institute of Crop Germplasm Resources, Shanxi Academy of Agricultural Sciences, Key Laboratory of Crop Gene Resources and Germplasm Enhancement on Loess Plateau, Ministry of Agriculture / Shanxi Key Laboratory of Genetic Resources and Genetic Improvement of Minor Crops, Taiyuan 030031, China
3 Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
4School of Life Sciences, Jilin University, Changchun 130012, China
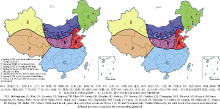
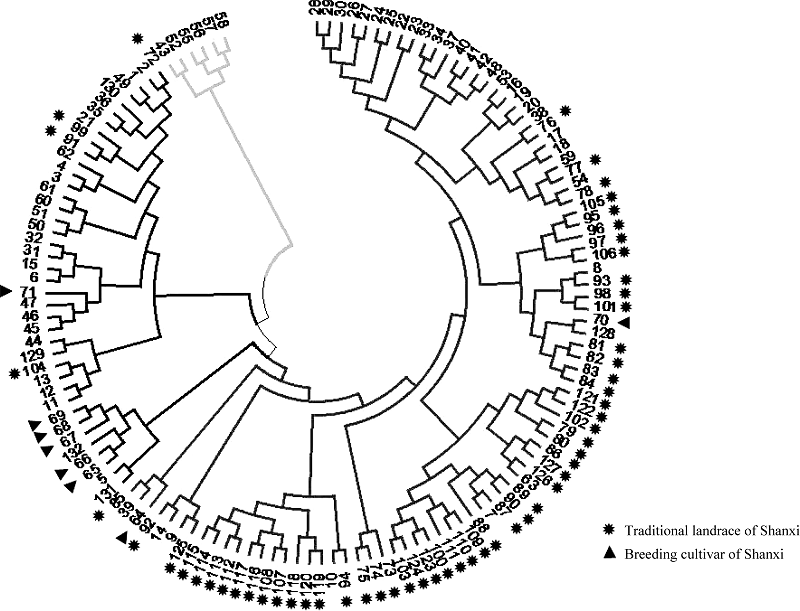
Fund:This study was supported by the National Natural Science Foundation of China (31271791), the Scientific Research Foundation for Returned Scholars in Shanxi province (2016-066), the National Natural Science Foundation of China for Young Scientists (31301386, 31300279), the China Agriculture Research System (CARS-07-13.5-A12), and General Project of the Shanxi Provincial Key Research and Development Program for Agriculture (201603D221003-5). AbstractUnderstanding of the genetic diversity of common millet germplasm resources is a basis of research on its origin and evolution, as well as the effective utilization of elite germplasm. In this study, we analyzed the genetic diversity of 132 common millet accessions collected from 11 provinces of China using 15 millet-specific fluorescent-labelled simple sequence repeat (SSR) markers. A total of 107 alleles were detected with 7 alleles per locus ranging from 2 to 14. The expected heterozygosity ranged from 0.0936 to 0.8676 with the mean value of 0.5298. The polymorphism information content (PIC) ranged from 0.0893 to 0.8538 with the mean value of 0.4864. The 132 accessions were clustered into four groups according to genetic distance between accessions, which showed geographic and eco-regional distributions. Group I is a collection of germplasm from spring-sowing area of Northeast China, group II is composed of partial accessions from the spring and summer-sowing areas of Loess Plateau, group III is composed of partial accessions from the spring-sowing area of Northern China, and the remaining accessions comprises Group IV with geographic overlaps to groups II and III. Model-based genetic structure analysis indicated that the genetic resources of Chinese common millet were from four gene pools in Northeast, Loess Plateau, Northern, and Northwest China. Obviously, this result was very similar to that of clustering analysis, showing the importance of geographical relationship. Rich genetic variations were also found in Chinese common millet resources, mainly from different accessions. The above results provide an overall evaluation of genetic dive rsity of common millet in China at a molecular level.
Keyword:Common millet ( Panicum miliaceum L.); Fluorescent SSR; Genetic diversity; Cluster analysis; Genetic structure Show Figures Show Figures
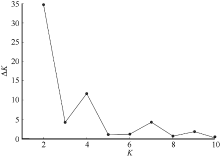
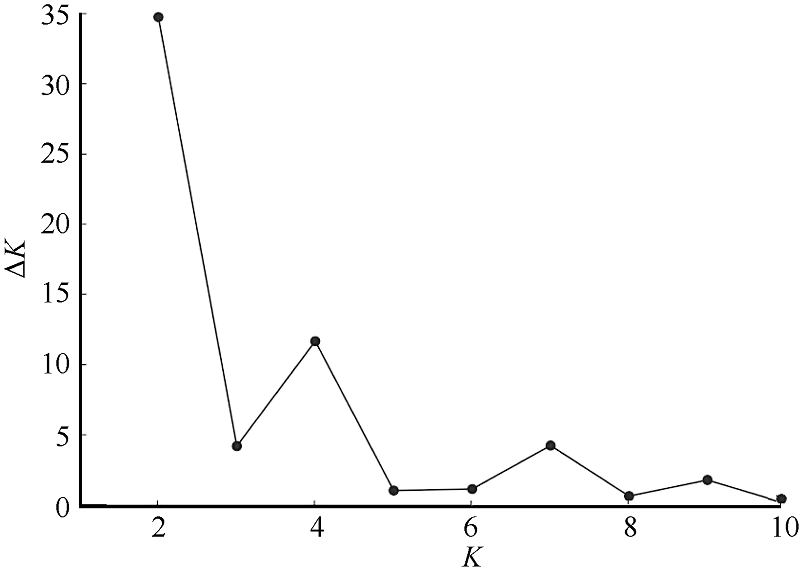
图2 运用Structure对糜子基因库数目建模 Delta K根据Evanno等[38]的方法计算得到, 针对基因库数目(K)建模。Fig. 2 Modelling of number of genepools in common millet using structure Delta K was calculated by the method of Evanno et al.[38] and model was established by the number of genepools (K).
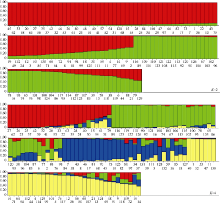
图3 糜子种质资源群体遗传结构图 横坐标的数字代表糜子材料序号。Fig. 3 Population genetic structure graph of accessions Numbers in the hori· zontal axis represent serial number of the accession.
LuH, ZhangJ, LiuK, WuN, LiY, ZhouK, YeM, ZhangT, ZhangH, YangX, ShenL, XuD, LiQ. Earliest domestication of common millet (Panicum miliaceum) in East Asia extended to 10000 years ago. Proc Natl Acad Sci USA, 2009, 106: 7367-7372[本文引用:1]
[2]
Martin JH, Leonard WH, Stamp DL. Principles of field crop production, 3rd edn. New York: MacMillan Publishing Co. , Inc. , 1976. pp 415-429[本文引用:1]
[3]
KurzevaA, RomanovaO, KrylovA. Genetic resources of common millet (Panicum miliaceum L. ) at the Vavilov Research Institute of Plant Industry (VIR). In: Chai Y, Feng B, eds. Advances in Broomcorn Millet Research: Proceedings of the 1st International Symposium on Broomcorn Millet. Yangling: Northwest A&F University Press, 2012. pp 76-81[本文引用:1]
[4]
王纶, 王星玉, 温琪汾, 武变娥. 中国黍稷种质资源研究与利用. 植物遗传资源学报, 2005, 6: 474-477WangL, Wang XY, Wen QF, Wu BE. Research and utilization of proso millet germplasm resource in China. J Plant Genet Resour, 2005, 6: 474-477 (in Chinese with English abstract)[本文引用:5]
[5]
Kobyzeva LN, Gorbacheva SN, Grigorashchenko LV, Gorlacheva OV, Biryukova OV. Formation of National collection and millet breeding in Ukraine. In: Chai Y, Feng B, eds. Advances in Broomcorn Millet Research: Proceedings of the 1st International Symposium on Broomcorn Millet. Yangling: Northwest A&F University Press, 2012. pp 67-73[本文引用:1]
[6]
LaglerR, GyulaiG, HumphreysM, SzaboZ, HorvathL, BittsanszkyA. Morphological and molecular analysis of common millet (P. miliaceum) cultivars compared to a DNA sample from the 15th century (Hungary). Euphytica, 2005, 146: 77-85[本文引用:2]
[7]
BagdiA, BalázsG, SchmidtJ, SzatmáriM, SchoenlechnerR, BerghoferE, TömösköziaS. Protein characterization and nutrient composition of Hungarian proso millet varieties and the effect of decortication. Acta Alimentaria, 2011, 40: 128-141[本文引用:1]
[8]
NishizawaN, FudamoY. The elevation of plasma concentration of high-density lipoprotein chloesterol in mice fed with protein from proso millet (Panicum miliaceum). Biosci Biotech Biochem, 1995, 59: 333-335[本文引用:1]
[9]
Park MY, Jang HH, Kim JB, Yoon HN, Lee JY, Lee YM, Kim JH, Park DS. Hog millet (Panicum miliaceum L. ): supplemented diet ameliorates hyperlipidemia and hepatic lipid accumulation in C57BL/6J-ob/obmice. Nutr Res Pract, 2011, 5: 511-519[本文引用:1]
[10]
刘峰. 糜子农艺性状的遗传多样性分析. 黑龙江农业科学, 2010, (3): 15-16LiuF. Genetic diversity analysis of agronomic traits on proso millet. Heilongjiang Agric Sci, 2010, (3): 15-16 (in Chinese with English abstract)[本文引用:2]
[11]
董孔军, 杨天育, 何继红. 甘肃省糜子地方资源农艺性状遗传多样性分析. 河北农业科学, 2012, 16(2): 1-3Dong KJ, Yang TY, He JH. Analysis of genetic diversity of agronomic traits of local broomcorn millet germplasm resources in Gansu province. Hebei Agric Sci, 2012, 16(2): 1-3 (in Chinese with English abstract)[本文引用:2]
[12]
胡兴雨, 陆平, 贺建波, 王纶, 王星玉, 张红生, 张宗文, 吴斌. 黍稷农艺性状的主成分分析与聚类分析. 植物遗传资源学报, 2008, 9: 492-496Hu XY, LuP, He JB, WangL, Wang XY, Zhang HS, Zhang ZW, WuB. Principal components and cluster analysis of agronomic traits of proso millet (Panicum miliaceum). J Plant Genet Resour, 2008, 9: 492-496 (in Chinese with English abstract)[本文引用:2]
[13]
胡兴雨, 王纶, 张宗文, 陆平, 张红生. 中国黍稷核心种质的构建. 中国农业科学, 2008, 41: 3489-3502Hu XY, WangL, Zhang ZW, LuP, Zhang HS. Establishment of broomcorn millet core collection in China. Sci Agric Sin, 2008, 41: 3489-3502 (in Chinese with English abstract)[本文引用:2]
[14]
董俊丽, 王海岗, 陈凌, 王君杰, 曹晓宁, 王纶, 乔治军. 糜子骨干种质资源遗传多样性和遗传结构分析. 中国农业科学, 2015, 48: 3121-3131Dong JL, Wang HG, ChenL, Wang JJ, Cao XN, WangL, Qiao ZJ. Analysis of genetic diversity and structure of proso millet core germplasm. Sci Agric Sin, 2015, 48: 3121-3131 (in Chinese with English abstract)[本文引用:12]
[15]
王瑞云, 季煦, 刘晓欢, 杨阳, 徐凤杰, 程冀南. 不同生态区糜子种质间叶片及光合特性的多样性分析. 山西农业大学学报(自然科学版), 2014, 34(2): 97-102Wang RY, JiX, Liu XH, YangY, Xu FJ, Cheng JN. Diversity of leaf traits and photosynthetic characteristics in broomcorn millet (Panicum miliaceum L. ) germplasms among different ecotype zone of China. J Shanxi Agric Univ (Nat Sci Edn), 2014, 34(2): 97-102 (in Chinese with English abstract)[本文引用:2]
[16]
王纶, 温琪汾, 曹利萍, 王星玉. 黍稷抗旱种质筛选及抗旱机理研究. 山西农业科学, 2007, 35(4): 31-34WangL, Wen QF, Cao LP, Wang XY. Drought-resistant germplasm screening and drought-resistant mechanism in proso millet. J Shanxi Agric Sci, 2007, 35(4): 31-34 (in Chinese with English abstract)[本文引用:2]
[17]
王海岗, 陈凌, 王君杰, 曹晓宁, 董俊丽, 王纶, 杨天育, 乔治军. 20份山西糜子种质资源抗旱性综合评价. 中国农学通报, 2014, 30(36): 115-119Wang HG, ChenL, Wang JJ, Cao XN, Dong JL, WangL, Yang TY, Qiao ZJ. Comprehensive assessment of drought resistance of 20 proso millet germplasm resources in whole growth period. Chin Agric Sci Bull, 2014, 30(36): 115-119 (in Chinese with English abstract)[本文引用:2]
[18]
王纶, 王星玉, 温琪汾, 武变娥, 曹利萍. 中国黍稷种质资源耐盐性鉴定. 植物遗传资源学报, 2007, 8: 426-429WangL, Wang XY, Wen QF, Wu BE, Cao LP. Identification of salt tolerance in Chinese proso millet germplasm. Plant Genet Resour, 2007, 8: 426-429 (in Chinese with English abstract)[本文引用:2]
[19]
刘敏轩, 张宗文, 吴斌, 陆平. 黍稷种质资源芽、苗期耐中性混合盐胁迫评价与耐盐生理机制研究. 中国农业科学, 2012, 45: 3733-3743Liu MX, Zhang ZW, WuB, LuP. Evaluation of mixed salt-tolerance at germination stage and seedling stage and the related physiological characteristics of Panicum miliaceum L. Sci Agric Sin, 2012, 45: 3733-3743 (in Chinese with English abstract)[本文引用:2]
[20]
王纶, 王星玉, 温琪汾, 赵卫红, 刘金玉. 中国黍稷种质资源抗黑穗病鉴定评价. 植物遗传资源学报, 2008, 9: 497-501WangL, Wang XY, Wen QF, Zhao WH, Liu JY. Identification and evaluation of resistance to dustbrand in Chinese proso millet germplasm resources. Plant Genet Resour, 2008, 9: 497-501 (in Chinese with English abstract)[本文引用:2]
[21]
Hu XY, Wang JF, LuP, Zhang HS. Assessment of genetic diversity in broomcorn millet (Panicum miliaceum L. ) using SSR markers. J Genet Genom, 2009, 36: 491-500[本文引用:5]
[22]
Rajput SG, HarvesonT, Santra DK. Development and characterization of SSR markers in proso millet (Panicum miliaceum L. ) based on switch grass genomics. Am J Plant Sci, 2014, 5: 175-186[本文引用:1]
[23]
Rajput SG, Santra DK. Evaluation of genetic diversity of proso millet (Panicum miliaceum) germplasm available in the USA using SSR markers. Crop Sci, 2016, doi: DOI:10.2135/cropsci2015.10.0644[本文引用:1]
[24]
Hunt HV, Campana MG, Lawes MC, Park YJ, Bower MA, Howe CJ, Jones MK. Genetic diversity and phylogeography of broomcorn millet (Panicum miliaceum L. ) across Eurasia. Mol Ecol, 2011, 20: 4756-4771[本文引用:8]
[25]
Liu MX, XuY, He JH, ZhangS, Wang YY, LuP. Genetic diversity and population structure of broomcorn millet (Panicum miliaceum L. ) cultivars and land races in China based on microsatellite markers. Int J Mol Sci, 2016, 17(3). pii: E370. doi: DOI:10.3390/ijms17030370[本文引用:7]
[26]
刘晓鑫, 谢传晓, 赵琦, 路明, 曲延英, 张滨, 梁业红, 张世煌. 基于SSR荧光标记技术的玉米群体混合样本基因频率分析方法. 中国农业科学, 2008, 41: 3991-3998Liu XX, Xie CX, ZhaoQ, LuM, Qu YY, ZhangB, Liang YH, Zhang SH. Establishment of fluorescent SSR technique on detecting allelic frequency in maize (Zea mays L. ) populations with bulk sampling strategy. Sci Agric Sin, 2008, 41: 3991-3998 (in Chinese with English abstract)[本文引用:1]
[27]
郝晨阳, 王兰芬, 贾继增, 董玉琛, 张学勇. SSR荧光标记和银染技术的比较分析. 作物学报, 2005, 31: 144-149Hao CY, Wang LF, Jia JZ, Dong YC, Zhang XY. Comparison of fluorescence and silver-staining detection systems of microsatellite markers. Acta Agron Sin, 2005, 31: 144-149 (in Chinese with English abstract)[本文引用:1]
[28]
李会勇, 王天宇, 黎裕, 石云素, 宋燕春, 陆平. TP-M13自动荧光检测法在高梁SSR基因型鉴定中的应用. 植物遗传资源学报, 2005, 6: 68-70Li HY, Wang TY, LiY, Shi YS, Song YC, LuP. Application of the TP-M13 automated fluorescent-lablled system of SSR genotyping in sorghum. J Plant Genet Resour, 2005, 6: 68-70 (in Chinese with English abstract)[本文引用:1]
Hu YG, ZhuJ, LiuF, ZhangZ, ChaiY, WeiningS. Genetic diversity among Chinese land races and cultivars of broomcorn millet (Panicum miliaceum) revealed by the polymerase chain reaction. Ann Appl Biol, 2008, 153: 357-364[本文引用:2]
[31]
EdwardsK, JohnstoneC, ThompsonC. A simple and rapid method for the preparation of plant genomic DNA for PCR analysis. Nucl Acids Res, 1991, 19: 1349[本文引用:1]
[32]
Cho YI, Chung JW, Lee GA, Ma KH, DixitA, Gwag JG, Park YJ. Development and characterization of twenty-five new polymorphic microsatellite markers in proso millet (Panicum miliaceum L. ). Genes & Genomics, 2010, 32: 267-273[本文引用:5]
[33]
LiuK, Muse SV. PowerMarker: integrated analysis environment for genetic marker data. Bioinformatics, 2005, 21: 2128-2129[本文引用:1]
[34]
Yeh FC, Boyle TJ. Population genetic analysis of co-dominant and dominant markers and quantitative traits. Belg J Bot, 1997, 129: 157[本文引用:1]
[35]
TamuraK, PetersonD, PetersonN, StecherG, NeiM, KumarS. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance and maximum parsimony methods. Mol Biol Evol, 2011, 28: 2731-2739[本文引用:2]
[36]
FalushD, StephensM, Pritchard JK. Inference of population structure using multilocus genotype data: linked loci and correlated allele frequencies. Genetics, 2003, 164: 1567-1587[本文引用:1]
[37]
PeakallR, Smouse PE. GenAlEx 6. 5: genetic analysis in Excel. Population genetic software for teaching and research-an update. Bioinformatics, 2012, 8: 2537-2539[本文引用:1]
[38]
EvannoG, RegnautS, GoudetJ. Detecting the number of cluster of individuals using the software structure: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1]
[39]
LiC, DongY, LiuM, LuP, LiW, WangY, CuiX, ZhouH, XuY. Ancient DNA analysis of Panicum miliaceum (broomcorn millet) from a Bronze Age cemetery in Xinjiang, China. Veget Hist Archaeobot, 2016, 25: 469-477[本文引用:1]
[40]
KaramD, WestraP, Nissen SJ, Ward SM, Figueiredo J E F. Assessment of silver-stained AFLP markers for studying DNA polymorphism in proso millet (Panicum miliaceum L. ). Brazil J Bot, 2006, 29: 609-615[本文引用:1]
[41]
KaramD, WestraP, Nissen SJ, Ward SM, Figueiredo J E F. Genetic diversity among proso millet (Panicum miliaceum) biotypes assessed by AFLP technique. Planta Daninha, 2004, 22: 167-174[本文引用:1]
[42]
M’Ribu HK, Hilu KW. Detection of interspecific and intraspecific variation in Panicum millets through rand om amplified polymorphic DNA. Theor Appl Genet, 1994, 88: 412-416[本文引用:1]
[43]
王银月, 刘敏轩, 陆平, 乔治军, 杨天育, 李海, 崔喜艳. 构建黍稷分子遗传图谱SSR引物的筛选. 作物杂志, 2014, (4): 32-38Wang YY, Liu MX, LuP, Qiao ZJ, Yang TY, LiH, Cui XY. SSR primer selection to construct molecular genetic map for proso millet. Crops, 2014, (4): 32-38 (in Chinese with English abstract)[本文引用:2]
[44]
Rajput SG, Santra DK, SchnableJ. Mapping QTLs for morpho-agronomic traits in proso millet (Panicum miliaceum L. ). Mol Breed, 2016, 36: 37, DOI: DOI:10.1007/s11032-016-0460-4[本文引用:1]
[45]
季煦, 王瑞云, 曳水瑛, 刘晓欢, 杨阳, 刘笑瑜, 连帅. 基于SSR的中国糜子遗传多样性分析. 山西农业大学学报(自然科学版), 2015, 35: 137-142JiX, Wang RY, Ye SY, Liu XH, YangY, Liu XY, LianS. The analysis of genetic diversity in broomcorn millet (Panicum miliaceum L. ) based on SSR markers. J Shanxi Agric Univ (Nat Sci Edn), 2015, 35: 137-142 (in Chinese with English abstract)[本文引用:2]
[46]
连帅, 王瑞云, 马跃敏, 刘笑瑜, 季煦. 不同生态区糜子种质资源的遗传多样性分析. 山西农业大学学报(自然科学版), 2015, 35: 225-231LianS, Wang RY, Ma YM, Liu XY, JiX. Genetic diversity of broomcorn millet (Panicum miliaceum L. ) germplasms of different ecotype zone of China. J Shanxi Agric Univ (Nat Sci Edn), 2015, 35: 225-231 (in Chinese with English abstract)[本文引用:2]
[47]
王纶, 王星玉, 温琪汾, 师颖. 山西是黍稷的起源和遗传多样性中心. 植物遗传资源学报, 2009, 10: 465-470WangL, Wang XY, Wen QF, ShiY. Shanxi province is the center of the origin and genetic diversity of proso millet. J Plant Genet Resour, 2009, 10: 465-470 (in Chinese with English abstract)[本文引用:1]
[48]
FrankhamR, Ballou JD, BriscoeD. Introduction to Conservation Genetics. Cambridge: Cambridge University Press, 2010. pp 309-336[本文引用:1]
 , 季煦
, 季煦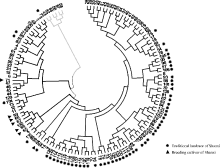






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}