关键词:水稻; splZ97; 斑点叶; 生理特性; 基因定位 Physiological Characters and Gene Mapping of a Spotted-leaf Mutant splZ97 in Rice WEI Li-Quan1,**, LUO Yan-Min1,**, WANG Wen-Qiang1, CHI Chang-Cheng1, HUANG Fu-Deng2, XIANG Xun1, CHENG Fang-Min1, PAN Gang1,* 1College of Agriculture and Biotechnology, Zhejiang University, Hangzhou 310058, China
2Institute of Crop and Nuclear Technology Utilization, Zhejiang Academy of Agricultural Sciences, Hangzhou 310021, China
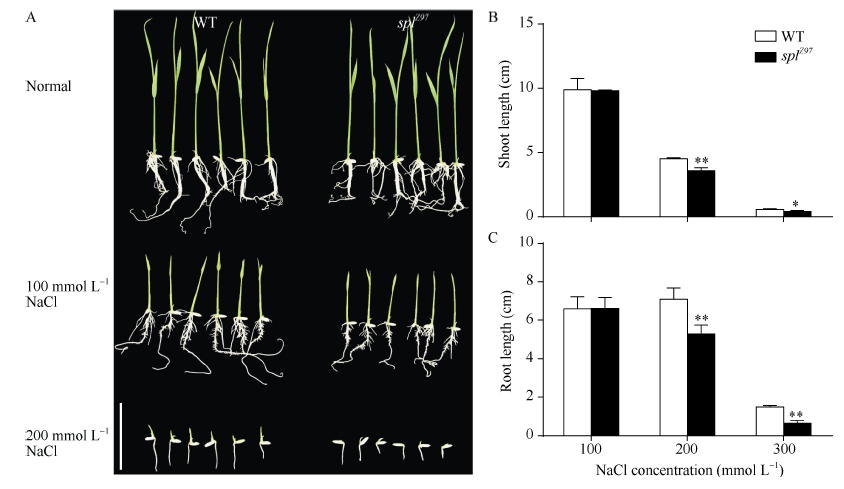
Fund:This study was supported by the Natural Science Foundation of China (31271691) and the National Major Project for Developing New GM Crops (2016ZX08001-002) AbstractA spotted-leaf mutant splZ97 was isolated from a mutant bank generated by EMS mutagenesis of indica restore line Zhen 97. Under field conditions, the brown lesion-mimics mutant splZ97 firstly displayed at the tip and edge of leaf blade at tillering stage, and then gradually spread to whole leaf, resulting in the death of the whole blade when the symptom was severe. At the same time, the major agronomic traits including plant height, grain number per panicle and seed-setting rate were markedly affected. Compared with the wild-type the flag leaf, the second leaf from top and the third leaf from top at heading stage, chlorophyll contents in the mutant splZ97 significantly decreased, while POD (peroxidase, POD) activity, O2? level and MDA (malondialdehyde, MDA) content increased. In addition, CAT (catalase, CAT) activity and soluble protein content of the second leaf from top and the third leaf from top of the mutant decreased as compared with the wild type; on the contrary, the SOD (superoxide dismutase, SOD) activity significantly increased. The histochemical analysis further indicated that O2? accumulated in the leaf blade of the mutant splZ97. In addition, under salt stress at seedling stage, the shoot length and root length of the mutant splZ97 were significantly shorter than these of the wild type. Genetic analysis and gene mapping showed that splZ97 was controlled by a single recessive nuclear gene, which was mapped to a region of 189 kb flanked by two SSR markers RM28466 and RM28485 on the long arm of chromosome 12. These results achieved in the present study would further facilitate the cloning and functional analysis of the gene SPLZ97.
Keyword:Rice; splZ97; Spotted-leaf; Physiological characters; Gene mapping Show Figures Show Figures
图1 突变体及其野生型的表型A: 分蘖期; B: 抽穗期; C: 抽穗期叶片; D: 开花期; E: 开花期叶片; F: 开花期倒二叶; 1~4代表剑叶至倒四叶。Bar=20 cm。Fig. 1 Phenotype of splZ97 and its wild-type (WT) plants at different growth stagesA: tillering stage; B: heading stage; C: leaves at heading stage; D: flowering stage; E: leaves at flowering stage; F: the 2nd leaf from top at the flowering stage; 1-4 mean the 1st leaf to 4th leaf from top. Bar=20 cm.
表1 Table 1 表1(Table 1)
表1 突变体及其野生型的农艺性状 Table 1 Agronomic traits ofsplZ97 and its wild-type (WT) plants
性状 Trait
2014
2015
野生型 WT
突变体 splZ97
野生型 WT
突变体 splZ97
株高 Plant height (cm)
111.23± 3.49
82.87± 3.48* *
104.11± 6.50
78.29± 4.99* *
穗长 Panicle length (cm)
23.28± 2.19
20.15± 1.97*
22.80± 1.34
20.04± 1.67*
有效穗数 Effective panicle number
9.45± 2.34
8.98± 1.57
9.78± 2.73
9.15± 0.95
每穗粒数 Grain number per panicle
125.58± 8.15
67.81± 5.67* *
130.06± 7.39
66.95± 6.49* *
结实率 Seed-setting rate (%)
81.34± 3.29
47.89± 2.48* *
85.83± 4.26
48.07± 4.32* *
千粒重 1000-grain weight (g)
26.92± 0.52
25.75± 0.61
26.60± 0.57
25.69± 0.71
* Significantly different at P < 0.05; * * significantly different at P < 0.01 (t-test). * 在0.05水平上差异显著; * * 在0.01水平上差异显著。
表1 突变体及其野生型的农艺性状 Table 1 Agronomic traits ofsplZ97 and its wild-type (WT) plants
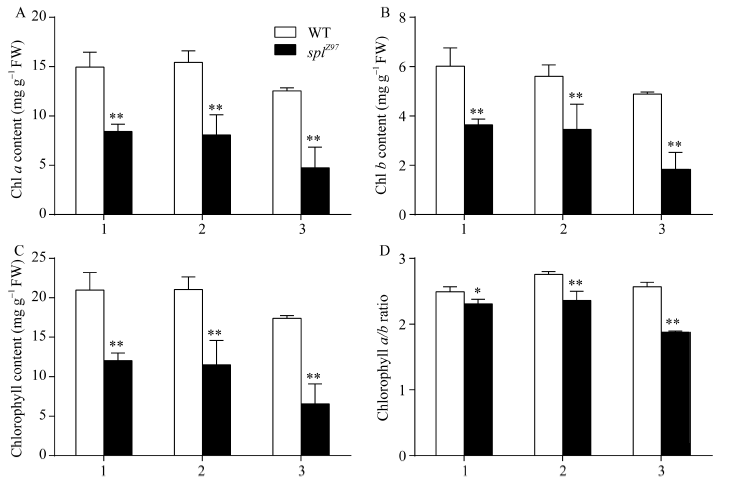
图3 孕穗期突变体及其野生型叶片的叶绿素含量1: 剑叶; 2: 倒二叶; 3: 倒三叶。* 在0.05水平上差异显著; * * 在0.01水平上差异显著。Fig. 3 Chlorophyll content of leaves in splZ97 and its wild-type (WT) plants at the booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * Significantly different at P< 0.05; * * significantly different at P< 0.01 (t-test).
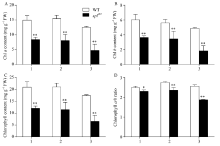
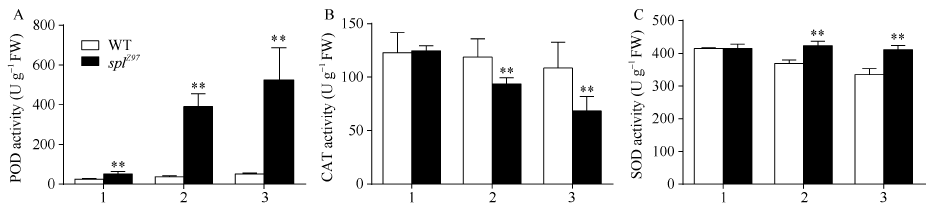
图4 孕穗期突变体splZ97及其野生型叶片的POD、CAT和SOD活性1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。Fig. 4 POD, CATs, and SOD activities of splZ97and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test).
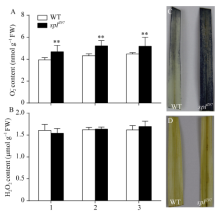
图5 孕穗期突变体splZ97及其野生型叶片的O2?和H2O2累积1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。C: NBT染色。D: DAB染色。Fig. 5 Accumulation of O2?and H2O2 in splZ97 and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test). C: NBT staining.D: DAB staining.
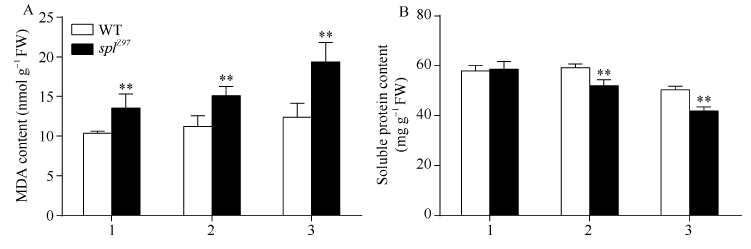
图6 孕穗期突变体splZ97及其野生型叶片的MDA和可溶性蛋白含量1: 剑叶; 2: 倒二叶; 3: 倒三叶。* * 在0.01水平上差异显著。Fig. 6 MDA and soluble protein contents of splZ97 and its wild-type (WT) plants at booting stage1: flag leaves; 2: 2nd leaves from top; 3: 3rd leaves from top. * * Significantly different at P< 0.01 (t-test).
孙惠敏, 张春娇, 李保同, 潘晓华. 水稻类病斑突变体的研究进展. 上海农业学报, 2014, 30: 142-147Sun HM, Zhang CQ, Li BT, Pan XH. Advances of study on rice lesion mimic mutants. Acta Agric Shanghai, 2014, 30: 142-147 (in Chinese with English abstract)[本文引用:1]
[2]
MoederW, YoshiokaK. Lesion mimic mutants: a classical, yet still fundamental approach to study programmed cell death. Plant Signal Behav, 2008, 3: 764-767[本文引用:2]
[3]
WuC, BordeosA, Madamba MR, BaraoidanM, RamosM, Wang GL, Leach JE, LeungH. Rice lesion mimic mutants with enhanced resistance to diseases. Mol Genet Genomics, 2008, 279: 605-619[本文引用:2]
[4]
SunC, LiuL, TangJ, LinA, ZhangF, FangJ, ZhangG, ChuC. RLIN1, encoding a putative coproporphyrinogen III oxidase, is involved in lesion initiation in rice. J Genet Genomics, 2011, 38: 29-37[本文引用:2]
[5]
ZhangH, CaoY, ZhaoJ, LiX, XiaoJ, WangS. A pair of orthologs of a leucine-rich repeat receptor kinase-like disease resistance gene family regulates rice response to raised temperature. BMC Plant Biol, 2011, 11: 160[本文引用:2]
[6]
Yang DL, YangY, HeZ. Roles of plant hormones and their interplay in rice immunity. Mol Plant, 2013, 6: 675-685[本文引用:1]
[7]
BruggemanQ, RaynaudC, BenhamedM, DelarueM. To die or not to die? Lessons from lesion mimic mutants. Front Plant Sci, 2015, 6: 24[本文引用:4]
[8]
LorrainS, VailleauF, BalaguéC, RobyD. Lesion mimic mutants: keys for deciphering cell death and defense pathways in plants. Trends Plant Sci, 2003, 8: 263-271. [本文引用:3]
[9]
YamanouchiU, YanoM, LinH, AshikariM, YamadaK. A rice spotted leaf gene, Spl7, encodes a heat stress transcription factor protein. Proc Natl Acad Sci USA, 2002, 99: 7530-7535[本文引用:2]
[10]
WangL, PeiZ, TianY, HeC. OsLSD1, a rice zinc finger protein, regulates programmed cell death and callus differentiation. Mol Plant Microbe Interact, 2005, 18: 375-384[本文引用:1]
[11]
Wang SH, Lim JH, Kim SS, Cho SH, Yoo SC, Koh HJ, SakurabaY, Paek NC. Mutation of SPOTTED LEAF3 (SPL3) impairs abscisic acid-responsive signaling and delays leaf senescence in rice. J Exp Bot, 2015, 66: 7045-7059. [本文引用:1]
[12]
JiangH, ChenY, LiM, XuX, WuG. Overexpression of SGR results in oxidative stress and lesion-mimic cell death in rice seedlings. J Integr Plant Biol, 2011, 53: 375-387[本文引用:1]
[13]
WangZ, WangY, HongX, HuD, LiuC, YangJ, LiY, HuangY, FengY, GongH, LiY, FangG, TangH, LiY. Functional inactivation of UDP-N-acetylglucosamine pyrophosphorylase 1 (UAP1) induces early leaf senescence and defense responses in rice. J Exp Bot, 2015, 66: 973-987[本文引用:1]
[14]
FujiwaraT, MaisonneuveS, IsshikiM, MizutaniM, ChenL, Wong HL, KawasakiT, ShimamotoK. Sekiguchi lesion gene encodes a cytochrome P450 monooxygenase that catalyzes conversion of tryptamine to serotonin in rice. J Biol Chem, 2010, 285: 11308-11313[本文引用:1]
[15]
Zeng LR, QuS, BordeosA, YangC, BaraoidanM, YanH, XieQ, Nahm BH, LeungH, Wang GL. Spotted leaf 11, a negative regulator of plant cell death and defense, encodes a U-box/ armadillo repeat protein endowed with E3 ubiquitin ligase activity. Plant Cell, 2004, 16: 2795-2808[本文引用:2]
[16]
MoriM, TomitaC, SugimotoK, HasegawaM, HayashiN, Dubouzet JG, OchiaiH, SekimotoH, HirochikaH, KikuchiS. Isolation and molecular characterization of a spotted leaf 18 mutant by modified activation-tagging in rice. Plant Mol Biol, 2007, 63: 847-860[本文引用:1]
[17]
Jiao BB, Wang JJ, Zhu XD, Zeng LJ, LiQ, He ZH. A novel protein RLS1 with NB-ARM domains is involved in chloroplast degradation during leaf senescence in rice. Mol Plant, 2012, 5: 205-217[本文引用:1]
[18]
ChenX, HaoL, PanJ, ZhengX, JiangG, JinY, GuZ, QianQ, ZhaiW, MaB. SPL5, a cell death and defense-related gene, encodes a putative splicing factor 3b subunit 3 (SF3b3) in rice. Mol Breed, 2012, 30: 939-949[本文引用:1]
[19]
TamiruM, TakagiH, AbeA, YokotaT, KanzakiH, OkamotoH, SaitohH, TakahashiH, FujisakiK, OikawaK, UemuraA, NatsumeS, JikumaruY, MatsuuraH, UmemuraK, Terry MJ, TerauchiR. A chloroplast-localized protein LESION AND LAMINA BENDING affects defense and growth responses in rice. New Phytol, 2016, 210: 1282-1297[本文引用:1]
[20]
YangX, GongP, LiK, HuangF, ChengF, PanG. A single cytosine deletion in the OsPLS1 gene encoding vacuolar-type H+-ATPase subunit A1 leads to premature leaf senescence and seed dormancy in rice. J Exp Bot, 2016, 67: 2761-2776[本文引用:1]
[21]
QiaoY, JiangW, LeeJ, ParkB, Choi MS, PiaoR, Woo MO, Roh JH, HanL, Paek NC, Seo HS, Koh HJ. SPL28 encodes a clathrin-associated adaptor protein complex 1, medium subunit micro 1 (AP1M1) and is responsible for spotted leaf and early senescence in rice (Oryza sativa). New Phytol, 2010, 185: 258-274[本文引用:1]
[22]
FekihR, TamiruM, KanzakiH, AbeA, YoshidaK, KanzakiE, SaitohH, TakagiH, NatsumeS, Undan JR, UndanJ, TerauchiR. The rice (Oryza sativa L. ) LESION MIMIC RESEMBLING, which encodes an AAA-type ATPase, is implicated in defense response. Mol Genet Genomics, 2015, 290: 611-622[本文引用:1]
[23]
Manosalva PM, BruceM, Leach JE. Rice 14-3-3 protein (GF14e) negatively affects cell death and disease resistance. Plant J, 2011, 68: 777-787[本文引用:1]
[24]
Undan JR, TamiruM, AbeA, YoshidaK, KosugiS, TakagiH, YoshidaK, KanzakiH, SaitohH, FekihR, SharmaS, UndanJ, YanoM, TerauchiR. Mutation in OsLMS, a gene encoding a protein with two double-strand ed RNA binding motifs, causes lesion mimic phenotype and early senescence in rice (Oryza sativa L. ). Genes Genet Syst, 2012, 87: 169-179[本文引用:1]
[25]
MurashigeT, SkoogF. A revised medium for rapid growth and bio assays with tobacco tissue cultures. Physiol Plant, 1962, 15: 473-497[本文引用:1]
[26]
张治安, 陈展宇. 植物生理学实验技术. 长春: 吉林大学出版社, 2008. p 7Zhang ZA, Chen ZY. Experiment Technology of Plant Physiology. Changchun: Jilin University Press, 2008. p 7 (in Chinese)[本文引用:1]
[27]
KariolaT, BraderG, LiJ, Palva ET. Chlorophyllase 1, a damage control enzyme, affects the balance between defense pathways in plants. Plant Cell, 2005, 17: 282-294[本文引用:1]
[28]
MahalingamR, JambunathanN, Gunjan SK, FaustinE, WengH, AyoubiP. Analysis of oxidative signalling induced by ozone in Arabidopsis thaliana. Plant Cell Environ, 2006, 29: 1357-1371[本文引用:1]
[29]
Shen YJ, JiangH, Jin JP, Zhang ZB, XiB, He YY, WangG, WangC, Qian LL, LiX, Yu QB, Liu HJ, Chen DH, Gao JH, HuangH, Shi TL, Yang ZN. Development of genome-wide DNA polymorphism database for map-based cloning of rice genes. Plant Physiol, 2004, 135: 1198-1205[本文引用:1]
[30]
PanaudO, ChenX, McCouch S R. Development of microsatellite markers and characterization of simple sequence length polymorphism (SSR) in rice (Oryza sativa L. ). Mol Gen Genet, 1996, 252: 597-607[本文引用:1]
[31]
伍泽堂. 超氧自由基与叶片衰老时叶绿素破坏的关系. 植物生理学通讯, 1991, 27: 277-279Wu ZT. Relationship between superoxide radical and destruction of chlorophyll during leaf senescence. Plant Physiol Commun, 1991, 27: 277-279 (in Chinese with English abstract)[本文引用:1]
[32]
WangF, WuW, WangD, YangW, SunJ, LiuD, ZhangA. Characterization and genetic analysis of a novel light-dependent lesion mimic mutant, lm3, showing adult-plant resistance to powdery mildew in common wheat. PLoS One, 2016, 11: e0155358[本文引用:1]
[33]
HidegE, KalaiT, Kos PB, AsadaK, HidegK. Singlet oxygen in plants: its significance and possible detection with double (fluorescent and spin) indicator reagents. Photochem Photobiol, 2006, 82: 1211-1218[本文引用:1]
[34]
Dietrich RA, Richberg MH, SchmidtR, DeanC, Dangl JL. A novel zinc finger protein is encoded by the Arabidopsis LSD1 gene and functions as a negative regulator of plant cell death. Cell, 1997, 88: 685-694[本文引用:1]
[35]
华春, 王仁雷. 杂交稻及其三系叶片衰老过程中SOD、CAT活性和MDA含量的变化. 西北植物学报, 2003, 23: 406-409HuaC, Wang RL. Changes of SOD and CAT activities and MDA content during senescence of hybrid rice and three lines leaves. Acta Bot Boreali-Occident Sin, 2003, 23: 406-409 (in Chinese with English abstract)[本文引用:1]
[36]
YouJ, ChanZ. ROS regulation during abiotic stress responses in crop plants. Front Plant Sci, 2015, 6: 1092[本文引用:1]
[37]
汪媛. 水稻叶片衰老过程生理变化及蛋白质降解与蛋白酶活性变化研究. 扬州大学硕士学位论文, 江苏扬州, 2010WangY. The Research of Physiological Changes, Protein Degradation and Protease Activity in the Process of Leaf Senescence in Rice. MS Thesis of Yangzhou University, Yangzhou, China, 2010 (in Chinese with English abstract)[本文引用:1]
[38]
赵晨晨, 黄福灯, 龚盼, 杨茜, 程方民, 潘刚. 水稻叶片早衰突变体osled的生理特征与基因定位. 作物学报, 2014, 40: 1946-1955Zhao CC, Huang FD, GongP, YangX, Cheng FM, PanG. Physiological characteristics and gene mapping of a leaf early-senescence mutant osled in rice. Acta Agron Sin, 2014, 40: 1946-1955 (in Chinese with English abstract)[本文引用:1]
[39]
LindemoseS, O’Shea C, Jensen M K, Skriver K. Structure, function and networks of transcription factors involved in abiotic stress response. Int J Mol Sci, 2013, 14: 5842-5878[本文引用:1]
[40]
ShaoH, WangH, TangX. NAC transcription factors in plant multiple abiotic stress responses: progress and prospects. Front Plant Sci, 2015, 6: 902[本文引用:1]
[41]
AmbawatS, SharmaP, Yadav NR, Yadav RC. MYB transcription factor genes as regulators for plant responses: an overview. Physiol Mol Biol Plants, 2013, 19: 307-321[本文引用:1]
[42]
BakshiM, OelmüllerR. WRKY transcription factors: Jack of many trades in plants. Plant Signal Behav, 2014, 9: e27700[本文引用:1]
[43]
RaffaeleS, RivasS, RobyD. An essential role for salicylic acid in AtMYB30-mediated control of the hypersensitive cell death program in Arabidopsis. FEBS Lett, 2006, 580: 3498-3504[本文引用:1]
[44]
VailleauF, DanielX, TronchetM, Montillet JL, TriantaphylidèsC, RobyD. A R2R3-MYB gene, AtMYB30, acts as a positive regulator of the hypersensitive cell death program in plants in response to pathogen attack. Proc Natl Acad Sci USA, 2002, 99: 10179-10184[本文引用:1]
[45]
Mall TK, DweikatI, Sato SJ, NeresianN, XuK, GeZ, WangD, ElthonT, ClementeT. Expression of the rice CDPK-7 in sorghum: molecular and phenotypic analyses. Plant Mol Biol, 2011, 75: 467-479[本文引用:1]
[46]
宋莉欣, 黄奇娜, 奉保华, 施勇烽, 张晓波, 徐霞, 王惠梅, 李小红, 赵宝华, 吴建利. 水稻斑点叶突变体spl21的鉴定与基因定位. 作物学报, 2015, 41: 1519-1528Song LX, Huang QN, Feng BH, Shi YF, Zhang XB, XuX, Wang HM, Li XH, Zhao BH, Wu JL. Characterization and gene mapping of a spotted-leaf mutant spl21 in rice (Oryza sativa L. ). Acta Agron Sin, 2015, 41: 1519-1528 (in Chinese with English abstract)[本文引用:1]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}