 , 金黎平, 李广存, 卞春松, 徐建飞, 胡军, 屈冬玉
, 金黎平, 李广存, 卞春松, 徐建飞, 胡军, 屈冬玉*通讯作者(Corresponding author): 金黎平, E-mail:jinliping@caas.cn 第一作者联系方式: E-mail:duanshaoguang@caas.cn
收稿日期:2016-10-18 接受日期:2017-01-21网络出版日期:2017-02-20基金:
摘要
关键词:马铃薯; 种质资源; 农艺性状; SSR; 遗传多样性
Genetic Diversity Analysis of Potato Varieties
DUAN Shao-Guang
, JIN Li-Ping, LI Guang-Cun, BIAN Chun-Song, XU Jian-Fei, HU Jun, QU Dong-YuFund:
Abstract
Keyword:Potato; Germplasm; Agronomic trait; SSR maker; Genetic diversity
Show Figures
Show Figures


马铃薯及其近缘种分布于美国南部到南美洲南部的广大地区[1, 2, 3], 拥有极其庞大的次级基因库, 倍性从二倍体到六倍体均有覆盖, 种质资源十分丰富[4]。我国初期的马铃薯资源引进、搜集始于20世纪30年代[5]。随着与国外合作的日益深入, 我国的马铃薯种质资源数量和种类得到了很大的改善。中国农业科学院蔬菜花卉研究所保存了1500余份马铃薯材料, 包括马铃薯审定品种、国外引进品种、育种品系、野生种和二倍体单株等种质资源, 基本可以代表我国整体的马铃薯种质资源组成。
当今作物育种工作因遗传基础狭窄已进入瓶颈, 从长远的角度来讲, 如果不改变这种局面, 作物会更易遭受病虫害的侵袭[6, 7]。农艺性状和分子标记是育种家获取不同马铃薯资源间遗传差异的两种途径。其中, 形态学水平上的变异是最易观察的, 对农艺性状的观察和分类在早期的马铃薯种质资源研究中是马铃薯分类学的主要研究方法之一。目前, 中国的《马铃薯DUS测试指南》仍以形态性状测试为主, 该书详细规定了用于品种鉴定的表现型及其观测时期。当前马铃薯育种以形态评价为主, 分子标记辅助选择仅在少数抗病性状上使用[8]。因此, 开展马铃薯种质资源表型性状亲缘分析的研究是必要的。
基于PCR的SSR标记以其数量丰富、共显性遗传、多态性丰富等优点现已被广泛应用于马铃薯的品种鉴定、遗传关系分类、核心种质资源搜集评价等领域。如段艳凤[9]利用11对SSR引物构建了217份中国育成马铃薯品种的指纹图谱, 并进行了遗传多样性分析。Haan等[10]利用15对SSR引物对比了989份来自秘鲁中部的地方品种和173份外迁地方品种, 共检测到173个等位位点, 其中129个为2个群体所共享, 占总数的74.6%。Sharmab等[11]结合使用SSR、RTN等3种DNA分子标记, 对47份马铃薯品种进行多态性分析比较, 表明3种标记均可区分47份品种。然而, 存在的问题或是上述研究所用的材料数量少, 或是对当前的马铃薯资源组成不具有系统的代表性。
本研究利用保存的包括国内育成品种以及从欧洲、北美和CIP等地区和国际研究机构引进的559份材料, 从表型性状聚类及分子标记检测等两方面系统研究, 了解中国现有马铃薯栽培种的遗传多样性, 为拓宽我国马铃薯育成品种遗传基础提供参考。
1 材料与方法1.1 试验材料共收集马铃薯品种材料559份, 包括中国育成品种239份, 来自北美(美国和加拿大)的马铃薯品种101份, 来自欧洲的育成品种75份, 来自国际马铃薯中心(CIP)的材料108份。选取小叶子、米拉、燕子、卡它丁、疫不加、早玫瑰、中薯18、DM1-3 516 R44等8个由系谱分析获得的遗传差异大且具有广泛代表性的品种进行SSR引物筛选。于河北省张北县试验基地种植, 每份材料5株, 株行距为30 cm × 75 cm。559份材料用于SSR标记分析; 16个表型性状较全的454份材料用于表型性状的亲缘分析。
1.2 田间性状调查田间调查16项性状, 包括叶片性状(复叶大小、小叶密集程度、小裂叶数量、叶片颜色深浅、小叶大小和小叶宽度), 花冠性状(花冠大小、花冠花青素和花冠形状), 块茎性状(薯形、表皮颜色、薯肉颜色和芽眼深浅), 块茎品质(比重和炸片颜色)和成熟期。参考《马铃薯DUS测试指南》的分级标准。表型性状数据经标准化处理后, 用SPSS11.0软件进行UPGMA聚类分析。
1.3 马铃薯基因组DNA的提取与扩增采摘新鲜的、未感病虫害的马铃薯幼嫩叶片, 采用改良的CTAB法提取基因组DNA。PCR扩增体系为10 μ L, 包括模板DNA 2 μ L、10× 缓冲液1 μ L、dNTPs (10 μ mol L-1 each) 0.2 μ L、上下游引物各0.2 μ L、Taq酶0.2 μ L、ddH2O 6.2 μ L。扩增程序为94℃预变性5 min; 94℃变性30 s, 退火30 s, 72℃延伸20 s, 30个循环; 72℃延伸7 min。用6%非变性聚丙烯酰胺凝胶电泳检测扩增产物, 并用硝酸银染色观察结果, 采用0, 1系统记录谱带位置, 观察某一扩增条带的有无, 有带记为1, 无带记为0。
1.4 SSR引物来源与筛选所用SSR引物共975对, 其中一部分来自于公开发表的文献, 另一部分根据已经完成的马铃薯全基因组序列设计开发。根据引物序列信息比照测序基因组数据对所有引物进行染色体定位, 选取能够定位到马铃薯12条染色体上的引物, 利用8份遗传差异大并具有广泛代表性的材料进行筛选, 选择谱带清晰、多态性较高的SSR引物进行后续的自然群体筛选。引物均由上海生工生物工程技术服务有限公司合成。
1.5 数据处理和分析根据SSR标记数据, 利用NTSYS 2.11软件, 采用Jaccard’ s相似系数计算两两品种间的遗传相似系数, 得到相似系数矩阵。利用NTSYS-pc 2.11软件分析遗传多样性。Simpson’ s多样性指数也称位点多态信息量(PIC), PIC=1-Σ fi2; 每个位点的有效等位基因数为Ne=1/Σ Pi2, fi为基因座位上第i个等位基因变异出现的频率。借助生物分析软件PowerMarker V 3.25构建临接系统进化树(Neighbor-joining Tree, NJ), 参数为缺省。
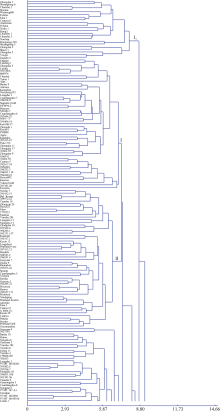
2 结果与分析2.1 基于表型性状的聚类分析根据454份国内外马铃薯材料16个表型性状的UPGMA聚类结果(图1), 在欧氏距离14.66处所有参试材料可以被聚成2个类群, 类群A1是一大类, 包括452份材料, 类群A只包括无花和甘农薯2号。在欧氏距离12.74处类群A1又可以被分为2个亚群A11和A12。在欧氏距离11.73处全部参试材料可以被聚成9个类群, 包括4个小类(A、B、C和H)和5个大类(D、E、F、G和I), 其中类群I所包括的材料占总数的57.5%。在欧氏距离8.65处该类群又可分为7个亚类(I1~I7), 其中较大的亚类I2在欧氏距离7.34处又可以进一步分为7个组(I~VII)。引自国际马铃薯中心的105份材料中的10份被集体聚于类群E, 35份被集体聚于类群D, 而且被聚于这2个类群内的材料距离都很近, 49份被集体聚于类群I, 且多数材料在数量上呈现两两及以上扎堆存在的局面, 且来源于国际马铃薯中心引进材料的合作系列品种(合作001、合作001、合作003和合作23)与国际马铃薯中心(CIP)材料的距离也较近; 而类群F以西南品种为主, 中原二作区等早熟品种主要聚在类群I。结果表明聚类分析可以较为准确地揭示马铃薯不同材料之间的形态差异, 进而区分生态类型不同和遗传差异明显的亲本。
图1-1
Fig. 1-1
| Figure OptionViewDownloadNew Window | |
 | 图1-1 参试454份马铃薯材料基于16个表型性状的聚类结果Fig. 1-1 Dendrogram of 454 materials based on 16 phenotype traits |
图1-2
Fig. 1-2
| Figure OptionViewDownloadNew Window | |
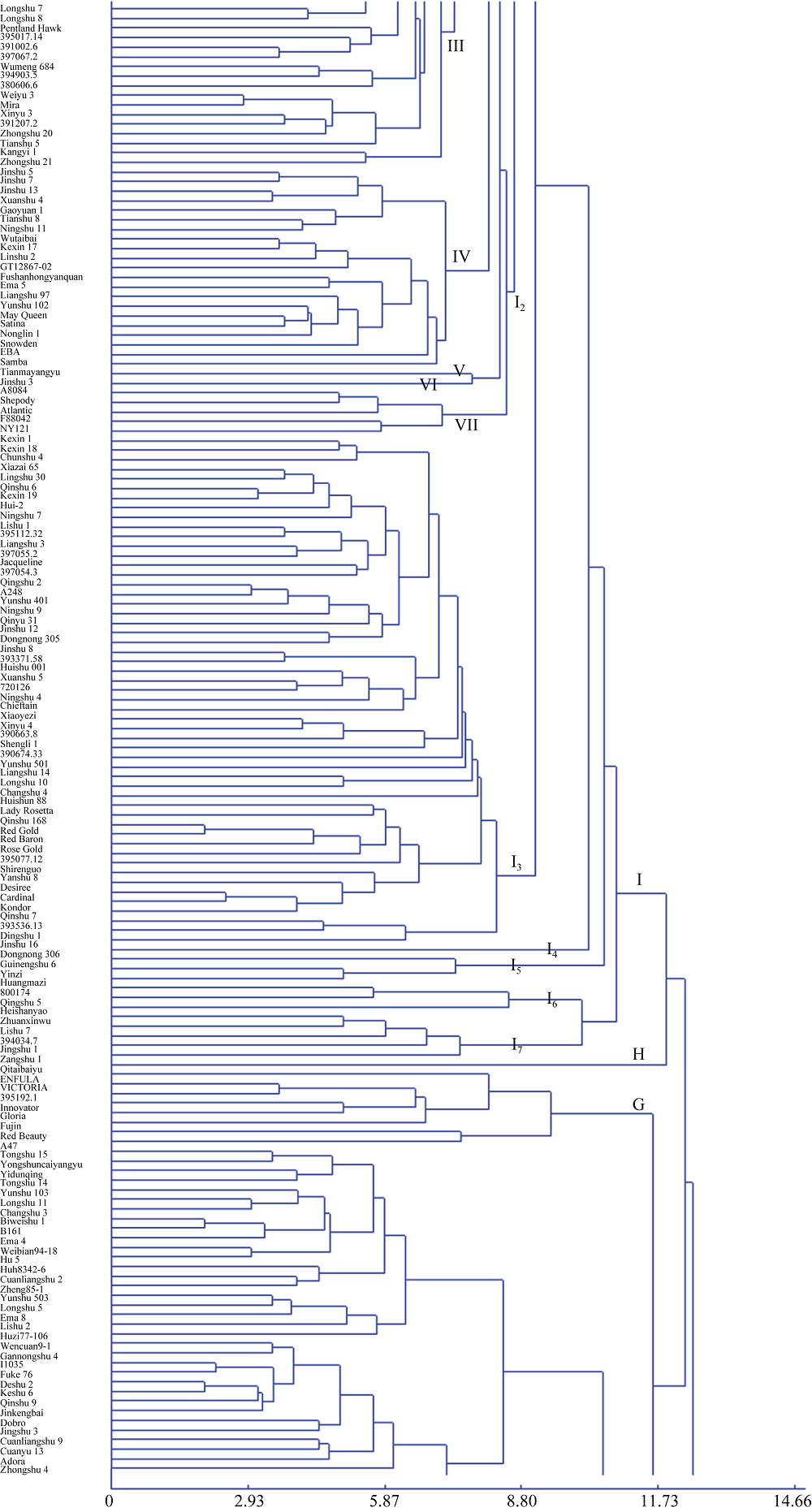 | 图1-2 参试454份马铃薯材料基于16个表型性状的聚类结果Fig. 1-2 Dendrogram of 454 materials based on 16 phenotype traits |
图1-3
Fig. 1-3
| Figure OptionViewDownloadNew Window | |
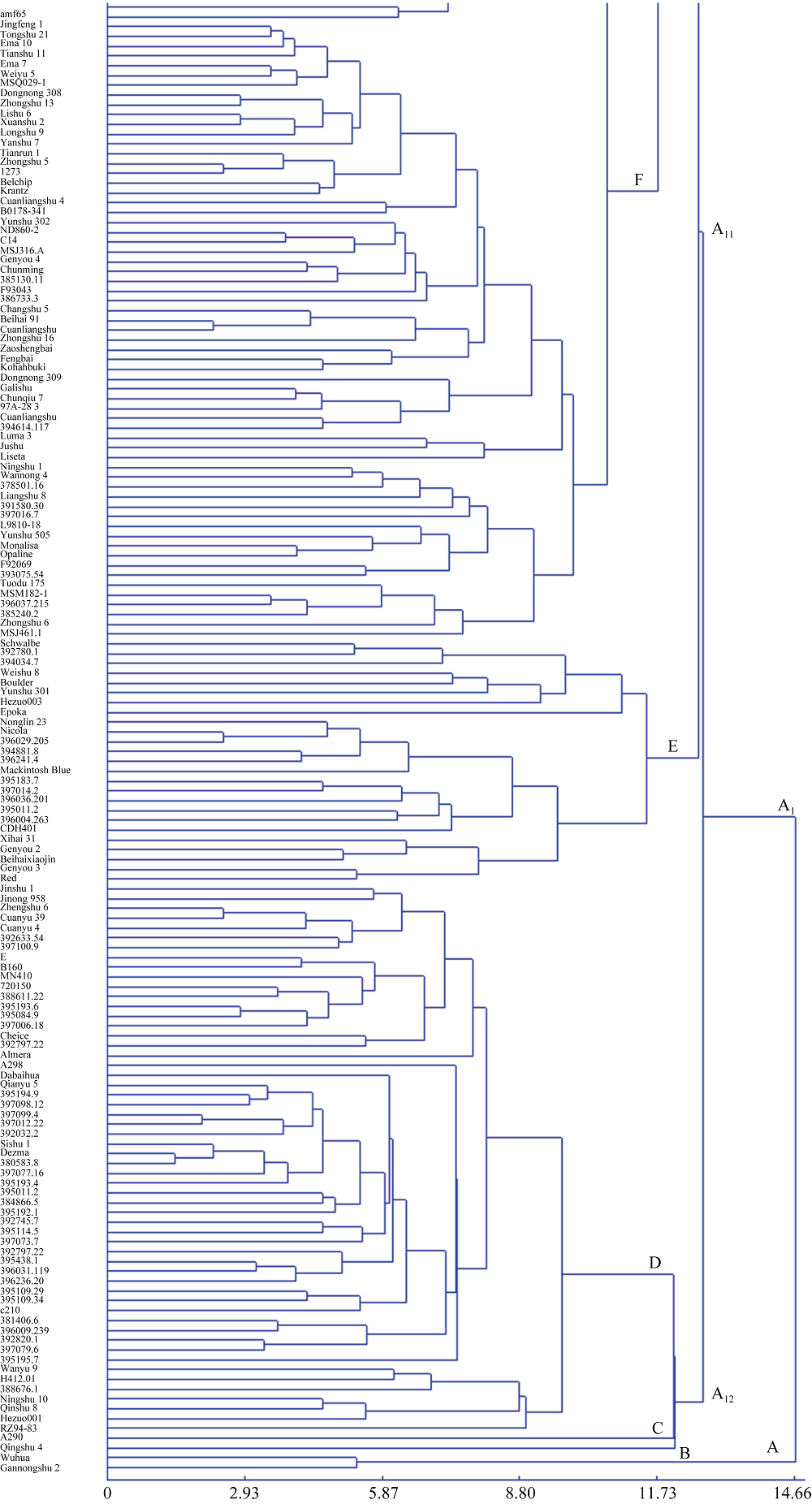 | 图1-3 参试454份马铃薯材料基于16个表型性状的聚类结果Fig. 1-3 Dendrogram of 454 materials based on 16 phenotype traits |
2.2 SSR标记多态性分析经过对SSR引物的比对, 975对引物中的332对成功地被定位到马铃薯的12条染色体上。采用染色体位置明确的332对SSR引物, 对遗传背景和田间性状差异较大的8个品种材料进行初步筛选, 得到36对扩增谱带清晰、条带差异明显的SSR引物, 36对引物均匀分布于马铃薯12条染色体上, 每条染色体上引物数量2~5对(表1)。
表1
Table 1
表1(Table 1)
| 表1 本研究所用SSR引物及其扩增结果 Table 1 SSR primers used in this study and their amplification results |

使用筛选出的36对SSR引物, 对559份马铃薯材料进行SSR分析。扩增结果显示, 36对引物共检测出134个等位变异, 引物等位位点数为1~7, 平均每对引物3.72个。其中, 引物PM0890有效等位基因数最多, 最少的为引物STM1049和SSR15406。36对引物扩增的产物大小为106~308 bp, 多态性信息量(PIC)的变化范围为0.1545~0.7743, 平均值为0.5783, 其中引物为SSR15406的PIC最低, 引物PM0890的PIC最高, PIC值所对应的引物和多态性位点所对应的引物是一致的, 可以反映较丰富的遗传多样性信息。
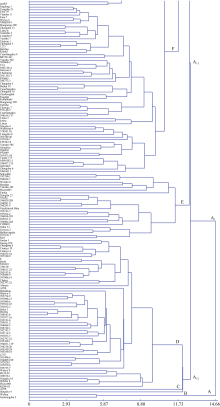
2.3 基于SSR标记的聚类分析图2所示, 559份不同来源的马铃薯品种总体分为3个大群, 类群I是一个混合群, 各地区品种均有分布, 包括133份马铃薯材料, 占总数的23.8%; 类群II中欧洲、北美以及中国东北和西北地区的材料所占比重大, 该类群包括的材料数量为187, 所占比例为33.5%; 类群III中北美、南美以及中国东北和西南地区马铃薯材料所占比重大, 其包含的239份材料占供试559份材料的42.8%。以上3个大的类群又可以细分为数量不同的小类群: 类群I可以再分为类群I-1和类群I-2, 类群II可以再分为类群II-1、类群II-2和类群II-3, 类群III可以再分为类群III-1、类群III-2和类群III-3。
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
 | 图2 供试559份材料的SSR临接系统进化树△ : 北美; ▲: 欧洲; ▽: CIP; ▼: 其他; ○: 中国东北; ●: 中国华北; ◇: 中国西北; ◆: 中国西南; □: 中国冬作中原; ■: IVF。Fig. 2 Neighbor-Joining tree based on dissimilarity matrix of 559 potato accessions examined with 36 microsatellite primer pairs△ : North America; ▲: Europe; ▽: CIP; ▼: Others; ○: Northeast China; ●: North China; ◇: Northwest China; ◆: Southwest China; □: Winter cropping Central Plain; ■: IVF. |
类群I-1是一个小的混合群, 含有31份材料, 各地区材料占比比较均匀, 代表品种为鄂马铃薯4号; 类群I-2是一个大的混合群, 含有102份材料, 各地区材料占比比较均匀, 代表品种为燕子, 虎头、坝薯10号均聚在这一类。而中国的中原二作区和冬作区早熟品种多聚在这一类, 如费乌瑞它、郑薯2号、春秋7号、克新18等, 中国农业科学院蔬菜花卉研究所育成的中薯系列品种也多聚在这一类, 包括中薯1号、中薯5号、中薯6号、中薯9号、中薯17和中薯19。
类群II-1是北美加工类群, 包含45份材料, 代表品种为Belchip, 美国和加拿大的多份加工品种(系)均聚在这一类; 类群II-2以欧洲以及中国东北和西北的育成品种(系)为主, 包含91份材料, 世界范围内很多育成的重要品种均聚在这一类, 如早玫瑰、抗疫白、克疫、白头翁、克新1号、东农303、陇薯3号、青薯2号等; 类群II-3为南美和欧洲类群, 包含51份材料, 代表品种有米拉和Spunta等。
类群III-1是南美和北美类群, 这一群含有南美48份材料, 代表品种392633.54 (B3C1)和Snowden, 另外具有CIP血缘的冀张薯8号、中薯20、合作002、合作23、云薯401、鄂马铃薯5号、川凉薯5号等均聚在这一类; 类群III-2为北美、欧洲和南美类群, 这一群含有北美29份材料, 代表品种小叶子、夏波蒂、大西洋、Red Pontiac和欧洲的Pentland Dell; 类群III-3是中国的东北和西南类群, 包含63份材料, 代表品种有火玛、卡它丁/春薯2号、内薯7号、云薯101、凉薯30, 以及一些老品种, 如圆叶青、一墩青、系薯1号等。
3 讨论3.1 马铃薯表型性状聚类分析的总体评价从本研究的结果来看, 表型性状的聚类分析可以较为准确地揭示马铃薯不同材料之间的形态差异, 进而区分生态类型不同和遗传差异明显的亲本, 这与徐敏[13]的研究结果相似。例如, 类群D中的材料绝大多数的成熟期很晚, 花冠的性状为近五边形, 小叶比较大; 类型E中材料的复叶较大、花冠大小中等、小裂叶数量多等。因此, 在马铃薯杂种优势的利用过程中, 对于杂交组合双亲表型性状不同的配制方式来说, 表型性状聚类分析具有一定的参考价值。
马铃薯的表型性状是由其遗传信息和生长环境条件共同决定的, 且易受后者的影响, 且马铃薯同源四倍体高度杂合的特性也决定了表型聚类分析只能从一定程度上反映不同材料的形态差异, 并不能从本质上体现马铃薯的遗传差异, 在本研究中得到了很好体现, 如云薯系列中的云薯101、云薯102、云薯201和云薯302是全同胞的关系, 均是杂交组合S95-105× 内薯7号的后代, 但这4个品种之间的距离并没有预期那样近, 前三者虽然均被聚在了I这一大类群的I2亚类群, 但却均不属于同一组(云薯101属于III组、云薯102属于IV组、云薯201属于II组), 而云薯302甚至没有被聚在类群I, 而是被聚在了距离很远的F组。此外, 考虑到可供调查的马铃薯表型性状很多, 不同的研究采用不同性状进行聚类分析时, 得到的结果也可能是不同的。从系谱来看, 威芋3号和凉薯14的亲缘关系很近, 徐敏[13]利用调查得到的20个表型性状的数据(部分表型性状与本研究不同), 确实将这2个品种聚在了一起, 而且距离很近(相邻), 但在本研究中, 2个品种却分别被聚在了类群I中的亚类群I2和亚类群I3, 由此可见, 该种聚类方法难以确定马铃薯品种间的遗传本质, 前人在其他作物的研究中也得到了类似的结果[14, 15]。另外, 对于诸如形状大小和颜色深浅等这样的性状, 调查者的主观因素比较大, 容易产生误差, 进而产生不同的聚类结果。因此, 表型性状聚类分析可以作为分子标记聚类分析的互补而发挥应有的作用。
3.2 SSR标记马铃薯资源聚类分析的总体评价考虑到多态性信息量(PIC)对于衡量SSR引物多态性高低的重要性, Botstein等[16]通过将PIC数值划分为3个区间PIC> 0.5, 0.25< PIC< 0.5和PIC< 0.25来对应引物多态性程度的高、中和低, 以此来衡量基因变异程度的高低。本研究从332对引物中筛选出36对, 其PIC值变幅为0.1545~0.7743, 其中引物SSR15406和STM1049的PIC小于0.25, 为低度多态性信息引物, STM1058、STG0026、PM0938、STM2022、STM1106、PM0355和PM0678等7对引物PIC值在0.25~0.50之间, 属于中度多态性信息引物, 其余27对引物的PIC值均大于0.5, 属于高度多态性信息引物, 比例占到了75%, 表明本研究所筛选得到的36对引物能较高信息量地反映中国马铃薯资源整体的基因型多样性水平, 再加上这些引物均匀地分布于马铃薯基因组的12条染色体, 反映了其整套染色体的遗传信息[16], 因此, 该36条SSR引物可以作为系统分析马铃薯种质资源遗传多样性的有效分子标记。作为对比, Ghislain等[17]采用156对SSR引物对属于8个分类群的930份马铃薯栽培种材料进行了遗传多样性分析, 其PIC的变幅为0.280~0.892。即是以PIC值这单一指标粗略对比, 也能够说明四倍体马铃薯栽培种的基因池并没有因为现代育种的影响而变窄, 这在之前的研究中也有提及[18]。
结合系谱分析和SSR分子标记分析, 除了可以明显区分马铃薯栽培种和野生种, 同时还可以将多数具有血缘关系的材料划分到一个类群。同一系列的品种(中薯系列多数分布在I-2类群、克新系列多数分布在II-2类群、云薯系列多数分布于III-3类群)亦趋向于分布在同一类群, 这说明各育种单位对某些具有优良祖先亲本的品种利用频率过于集中, 造成育成的品种亲缘关系较近, 遗传基础狭窄, 应加强品种亲本的多样性。当然, 这其中也存在一些品种聚类与已知系谱数据不一致, 如中薯5号是从中薯3号实生种子选育而来, 但二者并没有在同一类群, 这可能是由于36对SSR引物位点对于整个基因组而言很有限, 应用更多的标记将会减少这种不一致性, 同时这还可能是由马铃薯栽培品种之间的遗传基础狭窄造成的[19, 20, 21]。相对于传统的系谱分析[19, 22, 23], 应用分子标记来估计遗传关系将为马铃薯育种提供更多的信息。
本研究中, 按照材料的来源我们将材料区分成了北美、欧洲、CIP和中国国内等4个主要的地区, 参照中国马铃薯品种审定的区域试验地域的划分及育种单位的分布, 将中国材料的来源又分为东北、华北、西北、西南、冬作-中原和中国农业科学院蔬菜花卉研究所6个区域。SSR标记分析结果将所有材料分为了3个大类群, 又细分为8个小的类群。但我们发现除了欧洲地区之外, 所有区域内的材料基本是在1~2个大类群内或是1~4个小类群内扎堆出现, 基本没有在8个类群较均匀分布的情况, 这也说明马铃薯资源的遗传基础相对狭窄。马铃薯育种中应注重利用具有遗传多样性丰富的种质资源, 并在亲本选配时选择遗传距离较远且综合性状差异大的种质材料[24]。
3.3 表型性状聚类与SSR标记聚类结果比较本研究结果表明, 表型性状聚类与SSR分子标记聚类结果相似, 均与地理位置有很大相关性。例如来自CIP的类群D主要就聚在了类群III-1中, 来自中原二作区的早熟品种如早中薯5号、中薯6号、鲁马铃薯3号、德薯2号和春秋7号等表型聚类在类群F, SSR分子标记聚类在类群I-2; 但表型聚类与SSR聚类结果又有一定差异, 如同样多是来自CIP的材料在表型聚类中被聚在类群E, 在SSR聚类中却被分散在各个类群中。产生这种结果的原因可能是由于表型性状是基因表达和所处环境共同作用的结果, 表型聚类的原始数据随环境因素不同易出现变化, 且数据采集过程中人为因素也会产生误差, 因此表型性状不能完全真实反应物种的遗传多样性, 而SSR标记则是以基因组为研究对象, 不受环境影响, 相对稳定, 因此表型性状与分子标记的结果存在一定差异。但表型差异与遗传信息又具有一定的相关性, 可以结合共同用于评价马铃薯品种遗传多样性。
4 结论表型聚类准确地揭示了马铃薯不同品种间的形态差异, 区分了生态类型不同和遗传差异明显的亲本。SSR遗传多样性分析反映了品种间较丰富的遗传多样性。表型性状聚类与SSR分子标记聚类结果相似, 均与地理位置有很大相关性, 应结合共同用于评价马铃薯品种遗传多样性。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}