关键词:棉花; 机采相关性状; 关联分析; 等位变异 Association Analysis and Exploration of Elite Alleles of Mechanical Harvest- Related Traits with SSR Markers in Upland Cotton Cultivars ( Gossypium hirsutum L .) WANG Juan1,**, DONG Cheng-Guang1,2,**, LIU Li1, KONG Xian-Hui1, WANG Xu-Wen1, YU Yu1,* 1Cotton research Institute, Xinjiang Academy of Agricultural and Reclamation Science / Key Laboratory of China Northwestern Inland Region, Ministry of Agriculture, Shihezi 832000, China
2College of Agriculture, Xinjiang Agricultural University, Urumqi 830052, China
Fund:This study was supported by the Doctorial Found of Xinjiang Production and Construction Corps (2013BB001), the Key S&T Projects of; Xinjiang Production and Construction Corps (2016AA001-1), and the National Natural Science Foundation of China (31260340). AbstractCotton suitable for mechanical harvest should have higher requirement in traits, for example, shorter growth period, ideal plant type and high sensitivity to defoliant. A total of 214 pairs of SSR with high polymorphism and uniform distribution on whole genome were used to scan polymorphism in 118 cotton varieties with one or more mechanical harvest-related traits. Molecular marker data and six phenotypic traits were analyzed by the method of MLM (mixed linear model) in Tassel 5.0 on the basis of population structure, analysis loci with elite allelic variation and typical materials carrying elite alleles were identified based on phenotypic effect values. We detected 460 alleles and 905 genotypes. The average genetic diversity index was 0.5151, and the average polymorphic information content (PIC) per marker was 0.4587. Ninety-nine markers achieved the aforementioned average values accounted for 46.3% of the total markers, shows that the SSR markers have more allelic variance and higher genetic diversity. All the 118 cotton varieties were divided into four subgroups by analysis of population genetic structure. There was no corresponding relation between each kind of group of materials and the geographical source. A total of 124 loci ( P<0.05) and 20 loci ( P<0.01) associated with mechanical harvest-related traits were detected by association analysis, with explained variance ranging from 2.23% to 14.15% and from 4.84% to 14.15% respectively. Based on the results of this study, we identified 11 typical materials, including Xi 7, Jinken 9, Y11, Yumian 18, AY-4, K2, Chaoyang 2, DZ22, Zhongmiansuo 43, C2, and Guanrongchangzao B14. The elite alleles and resources can be useful for marker-assisted selection breeding.
Keyword:Cotton; Mechanical harvest-related traits; Associate on analysis; Allelic variation Show Figures Show Figures
图1 2013-2014年在石河子和库尔勒6个表型性状的Boxplot图(箱图两端表示其性状的极值范围; 点圈表示个别极值; 中间直线表示性状中位数。BJ2013、NJ2013、BJ2014、NJ2014分别代表北疆2013 (石河子2013年环境)、南疆2013 (库尔勒2013年环境)、北疆2014 (石河子2014年环境)、南疆2014 (库尔勒2014年环境)。)Fig. 1 Boxplot of six phenotypic traits in Shihezi and Kuerle from 2013 to 2014(Both ends of boxplot indicate the extreme range of traits; the circles of both ends of boxplot indicate single maximum and minimum values; the lines in the middle of boxplot indicate median. BJ2013, NJ2013, BJ2014, and NJ2014 represent Shihezi 2013 environment, Korla 2013 environment, Shihezi 2014 environment, and Korla 2014 environment, respectively.)
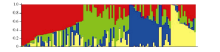
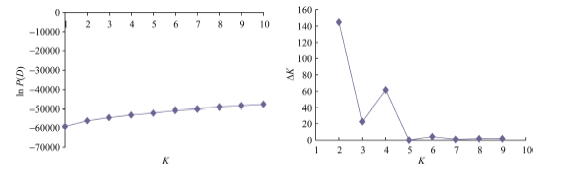
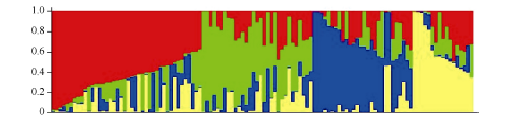
图2 118份材料的基于群体结构分析的K值与ln P(D)值和∆ K值变化图(A: K值与ln P(D)值的变化图; B: K值与∆ K值的变化图。)Fig. 2 ln P(D) and ∆ K based on population structure analysis for 118 upland cotton varieties(A: magnitude of ln P(D) as a function of K; B: magnitude of ∆ K as a function of K.)
中国农业科学院棉花研究所. 中国棉花遗传育种学. 济南: 山东科学技术出版社, 2003. p 562Cotton ResearchInstitute, the Chinese Academy of Agricultural Sciences. Genetics and Breeding of Cotton in China. Jinan: Shand ong Science&Technology Press, 2003. p 562 (in Chinese)[本文引用:1]
[2]
Guo WZ, Cai CP, Wang CB, Han ZG, Song XL, WangK, Niu XW, WangC, Lu KY, ShiB, Zhang TZ. A microsatellite-based, gene-rich linkage map reveals genome structure, function and evolution in Gossypium. Genetics, 2007, 176: 527-541[本文引用:1]
[3]
ZhaoL, Lyu YD, Cai CP, Tong XC, Chen XD, ZhangW, DuH, Guo XH, Guo WZ. Toward allotetraploid cotton genome assembly: integration of a high-density molecular genetic linkage map with DNA sequence information. BMC Genomics, 2012, 13: 1-17[本文引用:2]
[4]
范术丽, 喻树迅, 宋美珍, 原日红. 短季棉早熟性的分子标记及QTL定位. 棉花学报, 2006, 18: 135-139Fan SL, Yu SX, Song MZ, Yuan RH. Construction of molecular linkage map and QTL mapping for earliness in short-season cotton. Cotton Sci, 2006, 18: 135-139 (in Chinese with English abstract)[本文引用:1]
[5]
张先亮, 王坤波, 宋国立, 刘方, 黎绍惠, 王春英, 张香娣, 王玉红. 陆地棉重组近交系“中G6”QTL的初步定位. 棉花学报, 2008, 20: 192-197Zhang XL, Wang KB, Song GL, LiuF, Li SH, Wang CY, Zhang XD, Wang YH. Primary QTL mapping of upland cotton RIL CRI-G6 by SSR marker. Cotton Sci, 2008, 20: 192-197 (in Chinese with English abstract)[本文引用:1]
[6]
ChenH, QianN, Guo WZ, Song QP, Li BC, Deng FJ, Dong CG, Zhang TZ. Using three selected overlapping RILs to fine-map the yield component QTL on Chro. D8 in upland cotton. Euphytica, 2010, 76: 321-329[本文引用:1]
[7]
Liu RZ, Ai NJ, Zhu XX, Liu FJ, Guo WZ, Zhang TZ. Genetic analysis of plant height using two immortalized populations of “CRI12×J8891” in Gossypium hirsutum L. Euphytica, 2014, 196: 51-61[本文引用:1]
[8]
Ning ZY, ChenH, Mei HX, Zhang TZ. Molecular tagging of QTLs for fiber quality and yield in the upland cotton cultivar Acala-Prema. Euphytica, 2014, 195: 143-156[本文引用:1]
[9]
努斯热提·吾斯曼, 喻树迅, 范术丽, 庞朝友. 机采棉主要农艺性状相关性分析和QTL定位. 新疆农业科学, 2012, 49: 791-795NusuratO, Yu SX, Fan SL, Pang CY. Correlation of agronomic characters and QTL mapping in mechanical harvest cotton ( Gossypum hirsutum L. ) . Xinjiang Agric Sci, 2012, 49: 791-795 (in Chinese with English abstract)[本文引用:1]
[10]
Andersen JR, ZeinI, WenzelG, KrützfeldtB, EderJ, OuzunovaM, LübberstedtT. High levels of linkage disequilibrium and associations with forage quality at phenylalanine ammonia-lyase locus in European maize ( Zea mays L. ) inbreds. Theor Appl Gent, 2007, 114: 307-319[本文引用:1]
WangJ, McClean P E, Lee R, Goos R J, Helms T. Association mapping of iron deficiency chlorosis loci in soybean ( Glycine max L. Merr. ) advanced breeding lines. Theor Appl Genet, 2008, 116: 777-787[本文引用:1]
[13]
NiuY, XuY, Liu XF, Yang SX, Wei SP, Xie FT, Zhang YM. Association mapping for seed size and shape traits in soybean cultivars. Mol Breed, 2013, 31: 785-794[本文引用:1]
[14]
Patel JS, MamidiS, Bonman JM, Adhikari TB. Identification of QTL in spring wheat associated with resistance to a novel isolate of Pyrenophora tritici- repentis. Crop Sci, 2013, 53: 842-852[本文引用:1]
[15]
NarayananS, Vara Prasad P V, Shroyer K, Gill B S, Fritz A. Characterization of a spring wheat association mapping panel for root traits. Agron J, 2014, 106: 1593-1604[本文引用:1]
[16]
Malysheva-Otto LV, Ganal MW, Roder MS. Analysis of molecular diversity, population structure and linkage disequilibrium in a worldwide survey of cultivated barley germplasm( Hordeum vulgare L. ). BMC Genet, 2006, 7: 6[本文引用:1]
[17]
Liu GZ, Mei HX, WangS, Li XH, Zhu XF, Zhang TZ. Association mapping of seed oil and protein contents in upland cotton. Euphytica, 2015, 205: 637-645[本文引用:1]
[18]
Li CQ, Zhang JB, Hu GH, Fu YZ, Wang QL. Association mapping and favorable allele mining for node of first fruiting/sympodial branch and its height in upland cotton ( Gossypium hirsutum L. ). Euphytica, 2016, 210: 57-68[本文引用:1]
[19]
Li CQ, Ai NJ, Zhu YJ, Wang YQ, Chen XD, LiF, Hu QY, Wang Q L Association mapping and favorable allele exploration for plant architecture traits in Upland cotton ( Gossypium hirsutum L. ) accessions. J Agric Sci, 2016, 154: 567-583[本文引用:1]
[20]
聂新辉, 尤春源, 鲍健, 李晓方, 惠慧, 刘洪亮, 秦江鸿, 林忠旭. 基于关联分析的新陆早棉花品种农艺和纤维品质性状优异等位基因挖掘. 中国农业科学, 2015, 48: 2891-2910Nie XH, You CY, BaoJ, Li XF, HuiH, Liu HL, Qin JH, Lin ZX. Exploration of elite alleles of agronomic and fiber quality traits in Xinluzao cotton varieties by association analysis. Sci Agric Sin, 2015, 48: 2891-2910 (in Chinese with English abstract)[本文引用:1]
[21]
Mei HX, Zhu XF, Zhang TZ. Favorable QTL alleles for yield and its components identified by association mapping in Chinese upland cotton cultivars. PLoS One, 2013, 8: e82193[本文引用:1]
[22]
Qin HD, ChenM, Yi XD, BieS, ZhangC, Zhang YC, Lan JY, Meng YY, Yuan YL, Jiao CH. Identification of associated SSR markers for yield component and fiber quality traits based on frame map and Upland cotton collections. PLoS One, 2015, 10: e0118073[本文引用:2]
[23]
Cai CP, Ye WX, Zhang TZ, Guo WZ. Association analysis of fiber quality traits and exploration of elite alleles in upland cotton cultivars/accessions ( Gossypium hirsutum L. ). J Integr Plant Biol, 2014, 56: 51-62[本文引用:1]
[24]
Abdurakhmonov IY, Kohel RJ, Yu JZ, Pepper AE, Abdullaev AA, Kushanov FN, Salakhutdinov IB, Buriev ZT, SshaS, Scheffler BE. Molecular diversity and association mapping of fiber quality traits in exotic G. hirsutum L. germplasm. Genomics, 2008, 92: 478-487[本文引用:1]
[25]
ZhaoY, WangH, ChenW, Li YH. Genetic structure, linkage disequilibrium and association mapping of Verticillium wilt resistance in elite cotton ( Gossypium hirsutum L. ) germplasm population. PLoS One, 2014, 9: e86308[本文引用:1]
[26]
杜雄明, 周忠丽. 棉花种质资源描述规范和数据标准. 北京: 中国农业出版社, 2005Du XM, Zhou Z L. Descriptors and data stand ard for cotton (Gossypium spp. ). Beijing: China Agriculture Press, 2005 (in Chinese)[本文引用:1]
[27]
钱能. 陆地棉遗传多样性与育种目标性状基因(QTL)的关联分析. 南京农业大学博士学位论文, 江苏南京, 2009QianN. Genetic Diversity and Association of Gene (QTL) of Breeding Target Traits of Upland Cotton. PhD Dissertation of Nanjing Agricultural University, Nanjing, China, 2009 (in Chinese with English abstract)[本文引用:1]
[28]
Song XL, Zhang TZ. Quantitative trait loci controlling plant architectural traits in cotton. Plant Sci, 2009, 177: 317-323[本文引用:1]
[29]
薛艳, 张新宇, 沙红, 李雪源, 孙杰, 李保成. 新疆早熟棉品种SSR指纹图谱构建与品种鉴别. 棉花学报, 2010, 22: 360-366XueY, Zhang XY, ShaH, Li XY, SunJ, Li BC. Construction of fingerprinting map based on SSR and identification of cultivars for earliness cultivars in upland cotton in Xinjiang. Cotton Sci, 2010, 22: 360-366 (in Chinese with English abstract)[本文引用:1]
[30]
Sun FD, Zhang JH, Wang SF, Gong WK, Shi YZ, Liu AY, Li JW, Gong JW, Shang HH, Yuan YL. QTL mapping for fiber quality traits across multiple generations and environments in upland cotton. Mol Breed, 2012, 30: 569-582[本文引用:1]
[31]
艾先涛, 梁亚军, 沙红, 王俊铎, 郑巨云, 吐尔逊江, 多力坤, 李雪源, 华金平. 新疆自育陆地棉品种SSR遗传多样性分析. 作物学报, 2014, 40: 369-379Ai XT, Liang YJ, ShaH, Wang JZ, Zheng JY, Tu-Er-Xun-Jiang, Duo L K, Li X Y, Hua J P. Genetic diversity analysis on local upland cotton cultivars in Xinjiang based on SSR markers. Acta Agron Sin, 2014, 40: 369-379 (in Chinese with English abstract)[本文引用:1]
[32]
Paterson AH, Brubaker CL, Wendel JF. A rapid method for extraction of cotton ( Gossypium spp. ) genomic DNA suitable for RFLP and PCR analysis. Plant Mol Biol, 1993, 11: 122-127[本文引用:1]
[33]
张军, 武耀廷, 郭旺珍, 张天真. 棉花微卫星标记的PAGE/银染快速检测, 棉花学报, 2000, 12: 267-269ZhangJ, Wu YT, Guo WZ, Zhang TZ. Fast screening of microsatellite markers in cotton with PAGE/silver staining. Cotton Sci, 2000, 12: 267-269 (in Chinese with English abstract)[本文引用:1]
[34]
Liu KJ, Muse SV. PowerMarker: an integrated analysis environment for genetic marker analysis. Bioinformatics, 2005, 21: 2128-2129[本文引用:1]
EvannoG, RegnautsS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1]
[37]
Bradbury PJ, ZhangZ, Kroon DE, Casstevens TM, RamdossY, Buckler ES. TASSEL: software for association mapping of complex traits in diverse samples. Bioinformatics, 2007, 23: 2633-2635[本文引用:1]
[38]
Abdurakhmonov IY, SahaS, Jenkins JN, Buriev ZT, Shermatov SE, Scheffler BE, Pepper AE, Yu JZ, Kohel RJ, AbdukarimovA. Linkage disequilibrium based association mapping of fiber quality traits in G. hirsutumL. variety germplasm. Genetica, 2009, 136: 401-417[本文引用:1]
EvannoG, RegnautsS, GoudetJ. Detecting the number of clusters of individuals using the software STRUCTURE: a simulation study. Mol Ecol, 2005, 14: 2611-2620[本文引用:1]
[41]
Yang XH, Gao SB, Xu ST, Zhang ZX, Prasanna BM, LiL, Li JS, Yan JB. Characterization of a global germplasm collection and its potential utilization for analysis of complex quantitative traits in maize. Mol Breed, 2011, 28: 511-526[本文引用:1]
[42]
Pritchard JK, StephensM, Rosenberg NA, DonnellyP. Association mapping in structured populations. Am J Human Genet, 2000, 67: 170-181[本文引用:1]
[43]
ZhaoK, Aranzana MJ, KimS, ListerC, ShindoC, TangC, ToomajianC, ZhengH, DeanC, MarjoramP, NordborgM. An Arabidopsisexample of association mapping in structured samples. PLoS Genet, 2007, 3: 0071-0082[本文引用:1]
 , 董承光
, 董承光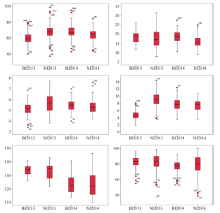



{kind=link}
{kind=link}
{kind=link}