关键词:棉花; 嫁接; 叶片; 衰老; 根-冠互作 Effect of Root-shoot Interaction on Cotton Leaf Senescence ZHANG Qiao-Yu, WANG Yi-Ru, AN Jing, WANG Bao-Min, TIAN Xiao-Li* Center of Crop Chemical Regulation, College of Agricultural and Biotechnology, China Agricultural University / State Key Laboratory of Plant Physiology and Biochemistry, Beijing 100193, China Fund:This study was supported by the National Natural Science Foundation of China (31271629) AbstractResearches and practices associated with grafting indicate that the diversity of root-shoot interaction is universal in different physiological processes of plant. Our previous grafting study with CCRI41, an early senescence cotton cultivar, and SCRC22, a late senescence cultivar as materials found that shoot played a major role in mediating leaf senescence of cotton. In the present study, CCRI41 and another late senescence cotton cultivar CCRI49 were used to do standard grafting (I-grafting, one scion grafted onto one rootstock) and Y-grafting (two scions grafted onto one rootstock). Leaf senescence of grafts was induced by low potassium (K; 0.03 mmol L-1). Contrary to the grafting combination of CCRI41 and SCRC22, the combination of CCRI41 and CCRI49 showed that the role of root was more important than that of shoot for leaf senescence, as characterized by SPAD reading. Accordingly, the concentration of ZR+Z or iPA+iP in roots and the youngest fully expanded leaf and their delivery rate in xylem sap regarding I-grafting combinations with CCRI41 as rootstock were significantly lower than those with CCRI49 as rootstock in most of situations. However, the results of free ABA level were exactly opposite. With respect to the Y-grafting of CCRI41 and CCRI49, the results were similar to those of I-grafting, whereas the role of root was not absolutely predominant any more. The mechanism for different root-shoot interactions in cotton leaf senescence was discussed from the perspective of root-shoot-root distance communication.
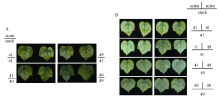
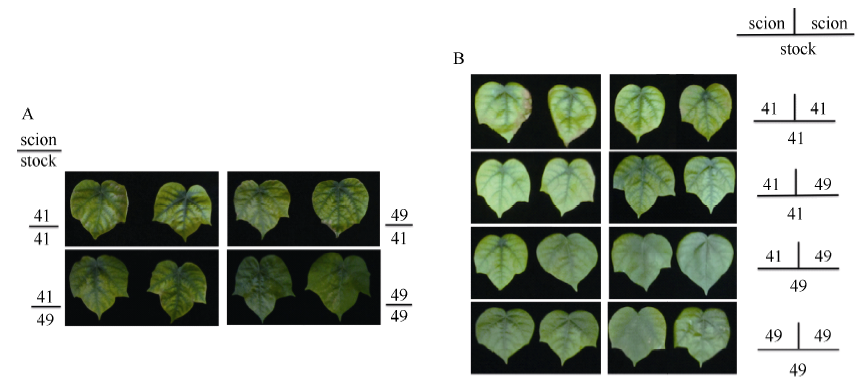
图1 高钾条件下(2.5 mmol L-1) I型(A)和Y型(B)嫁接植株的倒四叶Fig. 1 The youngest fully expanded leaf of I grafts (A) and Y grafts (B) under sufficient potassium (2.5 mmol L-1)
图2 低钾条件下(0.03 mmol L-1) I型(A)和Y型(B)嫁接植株的倒四叶Fig. 2 The youngest fully expanded leaf of I grafts (A) and Y grafts (B) under potassium deficiency (0.03 mmol L-1)
表1 Table 1 表1(Table 1)
表1 供钾水平对I型和Y型嫁接各处理倒四叶SPAD值的影响 Table 1 Effect of potassium (K) supply on SPAD value of the youngest fully expanded leaf of cotton standard grafts (scion/rootstock) and Y grafts (scion+scion/rootstock)
嫁接方式 Grafting method
供K水平 K supply (mmol L-1)
嫁接处理 Combination of scion and rootstock
I型 Scion/rootstock
41/41
49/41
41/49
49/49
2.50
37.9 a
38.1 a
38.0 a
38.4 a
0.03
18.4 d
20.3 c
21.5 b
24.3 a
Y型scion+scion/rootstock
41+41/41
41+41/41
41+49/41
41+49/41
41+49/49
41+49/49
49+49/49
49+49/49
2.50
36.0 a
35.2 a
33.7 a
34.4 a
34.9 a
35.5 a
35.7 a
34.3 a
0.03
18.3 b
19.3 b
18.2 b
21.1 ab
20.4 ab
25.5 a
22.7 ab
23.5 ab
41: CCRI41, early senescence under K deficiency; 49: CCRI49, late senescence under K deficiency. The data within the same grafting method followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test. 41: 中棉所41; 49: 中棉所49。每列数据为粗体接穗的结果; 各嫁接方式不同嫁接处理平均值后的不同字母表示在0.05水平差异显著。
表1 供钾水平对I型和Y型嫁接各处理倒四叶SPAD值的影响 Table 1 Effect of potassium (K) supply on SPAD value of the youngest fully expanded leaf of cotton standard grafts (scion/rootstock) and Y grafts (scion+scion/rootstock)
表2 低钾条件下(0.03 mmol L-1)不同嫁接方式倒四叶SPAD值的砧木和接穗调控效应 Table 2 Effects of rootstock and scion on SPAD value of the youngest fully expanded leaf under low K+ (0.03 mmol L-1) condition (%)
嫁接方式 Grafting method
中棉所41 CCRI 41
中棉所49 CCRI 49
砧木效应 Rootstock effect
接穗效应 Sion effect
砧木效应 Rootstock effect
接穗效应 Sion effect
I型 One scion/one rootstock
-16.3
-11.6
16.5
10.3
Y型 Two scions/one rootstock
-8.5
-0.6
8.4
4.8
表2 低钾条件下(0.03 mmol L-1)不同嫁接方式倒四叶SPAD值的砧木和接穗调控效应 Table 2 Effects of rootstock and scion on SPAD value of the youngest fully expanded leaf under low K+ (0.03 mmol L-1) condition (%)
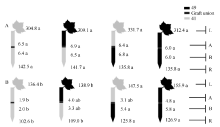
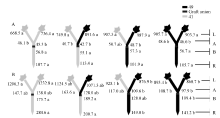
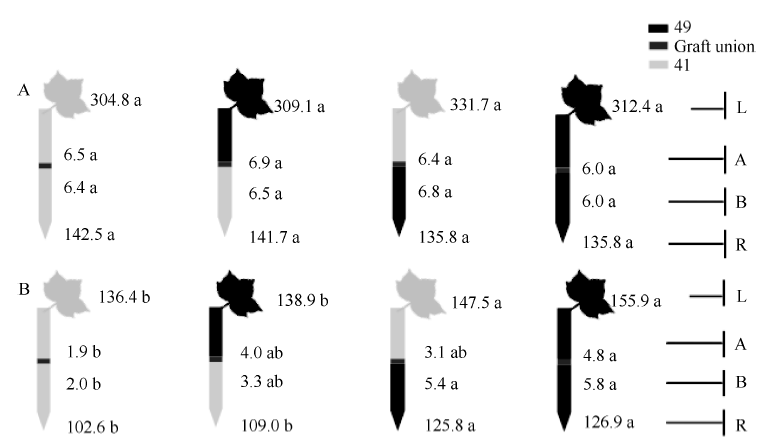
图3 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的ZR+Z浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ZR+Z流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 3 Effect of K deficiency on ZR+Z level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The ZR+Z concentration (ng g-1FW) in roots (R) and the youngest fully expanded leaf (L), and ZR+Z delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
图4 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的iPA+iP浓度(ng g-1 FW)及嫁接位点上(A)和下部(B)木质部汁液iPA+iP流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 4 Effect of K deficiency on iPA+iP level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The iPA+iP concentration (ng g-1FW) in roots (R) and the youngest fully expanded leaf (L), and iPA+iP delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
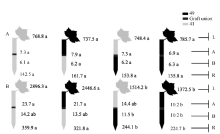
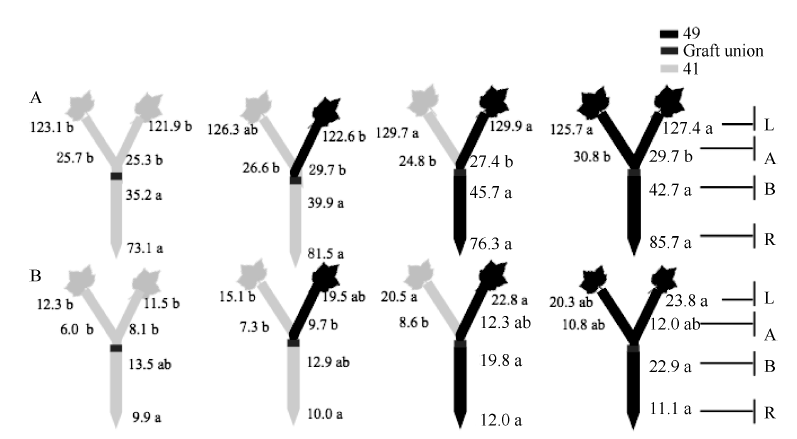
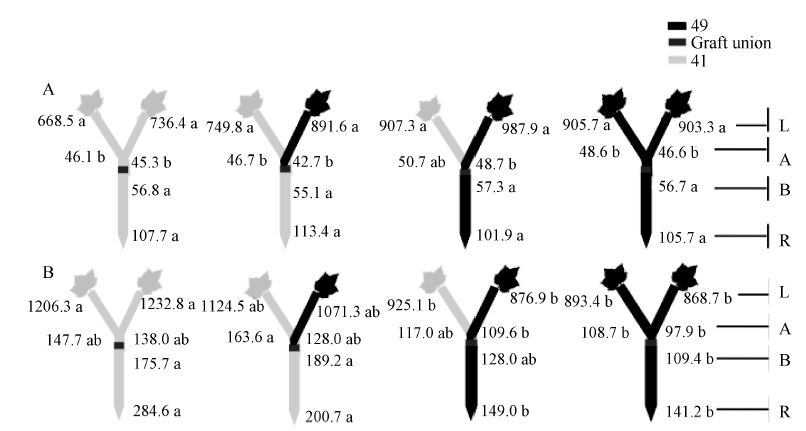
图5 供钾水平对I型嫁接各处理根系(R)和倒四叶(L)的ABA浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ABA流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 5 Effect of K deficiency on ABA level of cotton standard grafts (scion/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The ABA concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ABA delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
表3 低钾条件下(0.03 mmol L-1) I型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 3 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graftunion for standard graftings (one scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%)
植物激素 Phytohormone
中棉所41 CCRI 41
中棉所49 CCRI 49
砧木效应 Rootstock effect
接穗效应 Scion effect
砧木效应 Rootstock effect
接穗效应 Scion effect
倒四叶 The youngest fully expanded leaf
ZR+Z
-21.4
-9.2
25.9
9.1
iPA+iP
-10.9
-5.4
8.1
1.8
ABA
78.3
10.3
-47.7
-15.5
嫁接位点上部木质部汁液 The xylem sap above graft union
ZR+Z
-36.3
-17.3
50.0
15.6
iPA+iP
-36.0
-17.1
106.2
59.1
ABA
113.3
41.7
-39.1
-8.4
表3 低钾条件下(0.03 mmol L-1) I型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 3 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graftunion for standard graftings (one scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%)
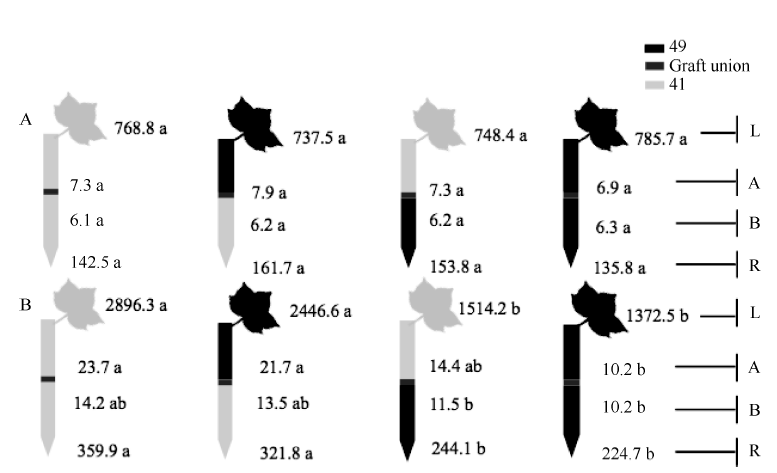
图6 供钾水平对Y型嫁接处理根系(R)和倒四叶(L)的ZR+Z浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ZR+Z流量 (ng plant-1 h-1)的影响 A:充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 6 Effect of K deficiency on ZR+Z level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1); B: low K (0.03 mmol L-1). The ZR+Z concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ZR+Z delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
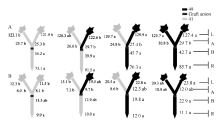
图7 供钾水平对Y型嫁接各处理根系(R)和倒四叶(L)的iPA+iP浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液iPA+iP流量(ng plant-1 h-1)的影响 A:充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 7 Effect of K deficiency on iPA+iP level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1). B: low K (0.03 mmol L-1). The iPA+iP concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and iPA+iP delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
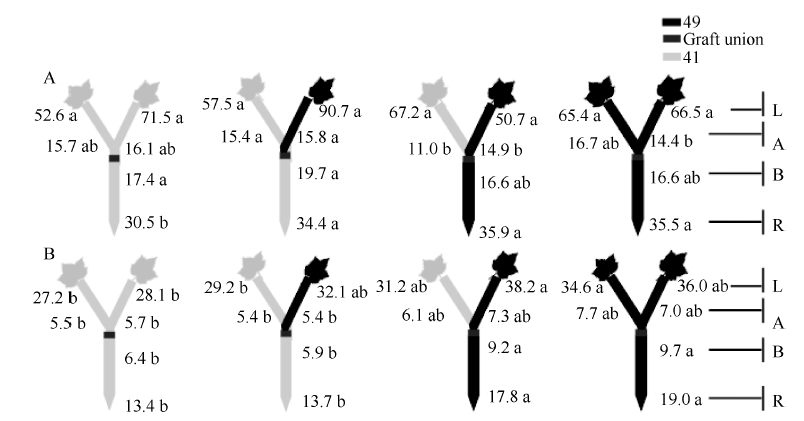
图8 供钾水平对Y型嫁接各处理根系(R)和倒四叶(L)的ABA浓度(ng g-1 FW)及嫁接位点上部(A)和下部(B)木质部汁液ABA流量(ng plant-1 h-1)的影响 A: 充足供钾(2.5 mmol L-1); B: 低钾胁迫(0.03 mmol L-1)。同一供钾水平, 同一部位(指根系、叶片和木质部汁液)数据后的不同小写字母表示在0.05水平差异显著(n = 4)。Fig. 8 Effect of K deficiency on ABA level of cotton Y grafts (two scions/rootstock) A: sufficient K (2.5 mmol L-1); B: low K (0.03 mmol L-1). The ABA concentration (ng g-1 FW) in roots (R) and the youngest fully expanded leaf (L), and ABA delivery rates (ng plant-124 h-1) in xylem above- (A) and below graft union (B) were determined. Means of the same sampling part (i.e. roots, leaf, and xylem sap collected both below and above the graft union) within the same K level followed by the same letter are not significantly different at P < 0.05 according to Duncan’ s multiple range test (n = 4).
表4 低钾条件下(0.03 mmol L-1) Y型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 4 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graf tunion for Y graftings (two scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%)
植物激素 Phytohormone
中棉所41 CCRI 41
中棉所49 CCRI 49
砧木效应 Rootstock effect
接穗效应 Scion effect
砧木效应 Rootstock effect
接穗效应 Scion effect
倒4叶 The youngest fully expanded leaf
ZR+Z
-11.7
-1.9
72.5
45.8
iPA+iP
-9.0
-1.7
13.0
10.8
ABA
21.6
2.3
-24.1
-10.0
嫁接位点上部木质部汁液 The xylem sap above graft union
ZR+Z
-15.2
-8.7
21.3
20.7
iPA+iP
-26.3
-8.6
8.4
4.2
ABA
23.9
9.7
-18.1
-2.1
表4 低钾条件下(0.03 mmol L-1) Y型嫁接倒四叶和接穗木质部汁液中激素水平的砧木和接穗效应 Table 4 Effect of rootstock and scion on phytohormone levels in the youngest fully expanded leaf and the xylem sap above graf tunion for Y graftings (two scion/one rootstock) under low K+ (0.03 mmol L-1) condition (%)
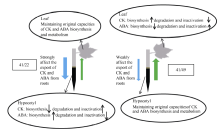
图9 棉花叶片衰老中不同根-冠互作类型的根-冠-根通讯机制 41: 中棉所41; 49: 中棉所49; 22: 鲁棉研22; 41/22: 地上部为主型; 41/49: 根系为主型; 绿色上行箭头: 根-冠信号; 蓝色下行箭头: 冠-根信号; 箭头粗细代表信号强弱。Fig. 9 The diagram of root-shoot-root communication for different interactions between root and shoot in cotton leaf senescence induced by low potassium (0.03 mmol L-1) stress 41: CCRI41; 49: CCRI49; 22: SCRC22; 41/22: the role of shoot is more important; 41/49: the role of root is more important. Green up arrow: the signal of root-shoot. Blue down arrow: the signal of shoot-root. The thickness of arrow indicates signal strength.
4 结论以中41为砧木的处理, 其叶片中ZR+Z、iPA+iP的含量低于以中49为砧木的处理, ABA含量则相反, 表明根系对叶片衰老的作用较大。据分析, 中41和中49互相嫁接体(41/49和49/41)接穗叶片中的受体感知到来自砧木的缺钾信号后, 发出的冠-根信号对砧木输出CK和ABA的能力和/或木质部中CK和ABA的运输过程影响较小; 接穗叶片中CK和ABA的合成、代谢则可能发生类似于砧木品种的变化。 致谢: 感谢课题组王晔博士在试验过程中提供的帮助。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
BreezeE, HarrisonE, McHattie S, Hughes L, Hickman R, Hill C, Kiddle S, Kim Y S, Penfold C A, Jenkins D. High-resolution temporal profiling of transcripts during Arabidopsis leaf senescence reveals a distinct chronology of processes and regulation. Plant Cell, 2011, 23: 873-894[本文引用:1]
[2]
FukaoT, YeungE, Bailey-SerresJ. The submergence tolerance gene SUB1A delays leaf senescence under prolonged darkness through hormonal regulation in rice. Plant Physiol, 2012, 160: 1795-1807[本文引用:1]
[3]
ChenY, DongH. Mechanisms and regulation of senescence and maturity performance in cotton. Field Crops Res, 2016, 189: 1-9[本文引用:1]
[4]
Kim HJ, RyuH, Hong SH, Woo HR, Lim PO, Lee IC, SheenJ, Nam HG, HwangI. Cytokinin-mediated control of leaf longevity by AHK3 through phosphorylation of ARR2 in Arabidopsis. Proc Natl Acad Sci USA, 2006, 103: 814-819[本文引用:2]
[5]
Gan SS, Amasino RM. Inhibition of leaf senescence by autoregulated production of cytokinin. Science, 1995, 270: 1986-1988[本文引用:3]
[6]
Lee IC, Hong SW, Whang SS, Lim PO, Nam HG, Koo JC. Age-dependent action of an ABA-inducible receptor kinase, RPK1, as a positive regulator of senescence in Arabidopsis leaves. Plant Cell Physiol, 2011, 52: 651-662[本文引用:2]
SongY, XiangF, ZhangG, MiaoY, MiaoC, Song CP. Abscisic acid as an internal integrator of multiple physiological processes modulates leaf senescence onset in Arabidopsis thaliana. Front Plant Sci, 2016, 7: 181[本文引用:2]
[9]
Letham DS. Zeatin, a factor inducing cell division isolated from Zea mays. Life Sci, 1963, 2: 569-573[本文引用:2]
AloniR, LanghansM, AloniE, DreieicherE, Ullrich CI. Root-synthesized cytokinin in Arabidopsis is distributed in the shoot by the transpiration stream. J Exp Bot, 2005, 56: 1535-1544[本文引用:3]
[12]
Rahayu YS, Walch-LiuP, NeumannG, Römheld V, von Wirén N, Bangerth F. Root-derived cytokinins as long-distance signals for NO3--induced stimulation of leaf growth. J Exp Bot, 2005, 56: 1143-1152[本文引用:2]
[13]
WernerT, MotykaV, LaucouV, SmetsR, Van OnckelenH, SchmüllingT. Cytokinin-deficient transgenic Arabidopsis plants show multiple developmental alterations indicating opposite functions of cytokinins in the regulation of shoot and root meristem activity. Plant Cell, 2003, 15: 2532-2550[本文引用:2]
[14]
KoD, KangJ, KibaT, ParkJ, KojimaM, DoJ, Kim KY, KwonM, EndlerA, Song WY. Arabidopsis ABCG14 is essential for the root-to-shoot translocation of cytokinin. Proc Natl Acad Sci USA, 2014, 111: 7150-7155[本文引用:3]
[15]
McKenzie MJ, MettV, Reynolds P HS, Jameson PE. Controlled cytokinin production in transgenic tobacco using a copper-inducible promoter. Plant Physiol, 1998, 116: 969-977[本文引用:1]
[16]
Letham DS, Palni L M S. The biosynthesis and metabolism of cytokinins. Annu Rev Plant Physiol Mol Biol, 1983, 34: 163-197[本文引用:1]
[17]
Hocart CH, Letham DS. Biosynthesis of cytokinin in germinating-seeds of Zea mays. J Expe Bot, 1990, 41: 1525-1528[本文引用:1]
[18]
WilkinsonS, Davies WJ. ABA-based chemical signalling: the co-ordination of responses to stress in plants. Plant Cell Environ, 2002, 25: 195-210[本文引用:2]
[19]
Dodd IC. Root-to-shoot signalling: assessing the roles of ‘up’ in the up and down world of long-distance signalling in planta. In: Lambers H, Colmer T D, eds. Root Physiology: from Gene to Function. Springer, 2005. pp 251-270[本文引用:2]
[20]
Dong HH, Niu YH, Li WJ, Zhang DM. Effects of cotton rootstock on endogenous cytokinins and abscisic acid in xylem sap and leaves in relation to leaf senescence. J Exp Bot, 2008, 59: 1295-1304[本文引用:1]
[21]
AlbaceteA, Martinez-AndujarC, Ghanem ME, AcostaM, Sanchez-BravoJ, Asins MJ, CuarteroJ, LuttsS, Dodd IC, Perez-AlfoceaF. Rootstock-mediated changes in xylem ionic and hormonal status are correlated with delayed leaf senescence, and increased leaf area and crop productivity in salinized tomato. Plant Cell Environ, 2009, 32: 928-938[本文引用:1]
[22]
Perez-AlfoceaF, AlbaceteA, Ghanem ME, Dodd IC. Hormonal regulation of source-sink relations to maintain crop productivity under salinity: a case study of root-to-shoot signalling in tomato. Funct Plant Biol, 2010, 37: 592-603[本文引用:1]
[23]
Ghanem ME, AlbaceteA, Smigocki AC, FrebortI, PospisilovaH, Martinez-AndujarC, AcostaM, Sanchez-BravoJ, LuttsS, Dodd IC, Perez-AlfoceaF. Root-synthesized cytokinins improve shoot growth and fruit yield in salinized tomato (Solanum lycopersicum L. ) plants. J Exp Bot, 2011, 62: 125-140[本文引用:1]
[24]
Beveridge CA, Murfet IC, KerhoasL, SottaB, MiginiacE, RameauC. The shoot controls zeatin riboside export from pea roots. Evidence from the branching mutant rms4. Plant J, 1997, 11: 339-345[本文引用:1]
[25]
FooE, Morris SE, ParmenterK, YoungN, WangH, JonesA, RameauC, Turnbull C G N, Beveridge C A. Feedback regulation of xylem cytokinin content is conserved in pea and Arabidopsis. Plant Physiol, 2007, 143: 1418-1428[本文引用:1]
[26]
FaissM, ZalubilovaJ, StrnadM, SchmullingT. Conditional transgenic expression of the ipt gene indicates a function for cytokinins in paracrine signaling in whole tobacco plants. Plant J, 1997, 12: 401-415[本文引用:1]
[27]
Koeslin-FindekleeF, Becker M A, van der Graaff E, Roitsch T, Horst W J. Differences between winter oilseed rape (Brassica napus L. ) cultivars in nitrogen starvation-induced leaf senescence are governed by leaf-inherent rather than root-derived signals. J Exp Bot, 2015, 66: 3669-3681[本文引用:2]
[28]
LiB, WangY, ZhangZ, WangB, Eneji AE, DuanL, LiZ, TianX. Cotton shoot plays a major role in mediating senescence induced by potassium deficiency. J Plant Physiol, 2012, 169: 327-335[本文引用:2]
[29]
WangY, LiB, Du MW, Eneji AE, Wang BM, Duan LS, Li ZH, Tian XL. Mechanism of phytohormone involvement in feedback regulation of cotton leaf senescence induced by potassium deficiency. J Exp Bot, 2012, 63: 5887-5901[本文引用:4]
[30]
熊长明, 王晔, 田晓莉. 植物矿质养分吸收的长距离反馈调节研究进展. 植物营养与肥料学报, 2014, 20: 737-746Xiong CM, WangY, Tian XL. Long-distance feedback regulation of mineral nutrients uptake in plant. J Plant Nutr Fert, 2014, 20: 737-746 (in Chinese with English abstract)[本文引用:1]
[31]
李博, 王春霞, 张志勇, 段留生, 李召虎, 田晓莉. 适用于低钾条件下棉花苗期根冠通讯研究的三种嫁接方法. 作物学报, 2009, 35: 363-369LiB, Wang CX, Zhang ZY, Duan LS, Li ZH, Tian XL. Three types of grafting techniques available for research of root-shoot communication in cotton (Gossypium hirsutum) seedlings under low-potassium condition. Acta Agron Sin, 2009, 35: 363-369 (in Chinese with English abstract)[本文引用:1]
[32]
AlexouM, Peuke AD. Methods for Xylem Sap Collection. In: Protocol: Plant Mineral Nutrients, Volume 953 of the Series Methods in Molecular Biology, Springer, 2013. pp 195-207[本文引用:1]
[33]
ZhaoJ, LiG, Yi GX, Wang BM, Deng AX, Nan TG, Li ZH, Li QX. Comparison between conventional indirect competitive enzyme-linked immunosorbent assay (icELISA) and simplified icELISA for small molecules. Anal Chim Acta, 2006, 571: 79-85[本文引用:1]
[34]
Estañ MT, Martinez-Rodriguez M M, Perez-Alfocea F, Flowers T J, Bolarin M C. Grafting raises the salt tolerance of tomato through limiting the transport of sodium and chloride to the shoot. J Exp Bot, 2005, 56: 703-712[本文引用:1]
[35]
SchwarzD, Öztekin GB, TüzelY, BrücknerB, KrumbeinA. Rootstocks can enhance tomato growth and quality characteristics at low potassium supply. Sci Hortic, 2013, 149: 70-79[本文引用:1]
[36]
PenellaC, Nebauer SG, QuiñonesA, San BautistaA, López- GalarzaS, CalatayudA. Some rootstocks improve pepper tolerance to mild salinity through ionic regulation. Plant Sci, 2015, 230: 12-22[本文引用:1]
[37]
朱进, 别之龙, 李娅娜. 黄瓜种子萌芽期及嫁接砧木幼苗期耐盐力评价. 中国农业科学, 2006, 39: 772-778ZhuJ, Bie ZL, Li YN. Evaluation of salt resistance of cucumber at seed germination and rootstock-seedling stages. Sci Agric Sin, 2006, 39: 772-778 (in Chinese with English abstract)[本文引用:1]
[38]
郝俊杰, 胡雨薇, 郭晓琴, 赵付安, 贾新合, 郭利娟, 张志新, 王庆东. 用相互嫁接和定量 PCR 分析棉花对棉花黄萎病的抗性. 作物学报, 2013, 39: 1179-1186Hao JJ, Hu YW, Guo XQ, Zhao FA, Jia XH, Guo LJ, Zhang ZX, Wang QD. Resistance to verticillium wilt in cotton by reciprocal grafting and real-time quantitative PCR. Acta Agron Sin, 2013, 39: 1179-1186 (in Chinese with English abstract)[本文引用:1]
[39]
阳燕娟, 王丽萍, 高攀, 郭世荣. 嫁接提高蔬菜作物抗逆性及其机制研究进展. 长江蔬菜, 2013, (22): 1-10Yang YJ, Wang LP, GaoP, Guo SR. Advances on mechanisms of improving stress resistance in rootstock-grafted vegetable crops. J Changjiang Veg, 2013, (22): 1-10 (in Chinese with English abstract)[本文引用:1]
[40]
高方胜, 王磊, 徐坤. 砧木与嫁接番茄产量品质关系的综合评价. 中国农业科学, 2013, 47: 605-612Gao FS, WangL, XuK. Comperhensive evaluation of relationship between rootstocks and yield and quality in grafting tomato. Sci Agric Sin, 2013, 47: 605-612 (in Chinese with English abstract)[本文引用:1]
[41]
ZhaoY. The role of local biosynthesis of auxin and cytokinin in plant development. Curr Opin Plant Biol, 2008, 11: 16-22[本文引用:1]
EmeryR, AtkinsC. Roots and cytokinins. In: Plant roots—the Hidden Half. New York: Marcel Dekker, 2002. pp 417-434[本文引用:1]
[44]
Kudoyarova GR, Korobova AV, Akhiyarova GR, Arkhipova TN, Zaytsev DY, PrinsenE, Egutkin NL, Medvedev SS, Veselov SY. Accumulation of cytokinins in roots and their export to the shoots of durum wheat plants treated with the protonophore carbonyl cyanide m-chlorophenylhydrazone (CCCP). J Exp Bot, 2014, 65: 2287-2294[本文引用:1]
[45]
BürkleL, CedzichA, DöpkeC, StranskyH, OkumotoS, GillissenB, KühnC, Frommer WB. Transport of cytokinins mediated by purine transporters of the PUP family expressed in phloem, hydathodes, and pollen of Arabidopsis. Plant J, 2003, 34: 13-26[本文引用:1]
[46]
HiroseN, MakitaN, YamayaT, SakakibaraH. Functional characterization and expression analysis of a gene, OsENT2, encoding an equilibrative nucleoside transporter in rice suggest a function in cytokinin transport. Plant Physiol, 2005, 138: 196-206[本文引用:1]
[47]
BrugiereN, Jiao SP, HantkeS, ZinselmeierC, Roessler JA, Niu XM, Jones RJ, Habben JE. Cytokinin oxidase gene expression in maize is localized to the vasculature, and is induced by cytokinins, abscisic acid, and abiotic stress. Plant Physiol, 2003, 132: 1228-1240[本文引用:1]
[48]
王逸茹. 根冠互作对棉花下胚轴细胞分裂素和脱落酸相关基因表达的影响. 中国农业大学硕士学位论文, 北京, 2015Wang YR. The Effect of Rootstock on the Expression of Cytokinin and Abscisic Acid Related Gene in Cotton Hypocotyl. MS Thesis of China Agricultural University, Beijing, China, 2015 (in Chinese with English abstract)[本文引用:1]
[49]
SinghS, Letham DS, Palni L M S. Cytokinin biochemistry in relation to leaf senescence: VIII. Translocation, metabolism and biosynthesis of cytokinins in relation to sequential leaf senescene of tobacco. Physiol Plant, 1992, 86: 398-406[本文引用:2]
[50]
SauterA, Dietz KJ, HartungW. A possible stress physiological role of abscisic acid conjugates in root-to-shoot signalling. Plant, Cell Environ, 2002, 25: 223-228[本文引用:1]
[51]
ZhangR, Letham DS, Willcocks DA. Movement to bark and metabolism of xylem cytokinins in stems of Lupinus angustifolius. Phytochemistry, 2002, 60: 483-488[本文引用:1]
[52]
Li BB, Feng ZG, XieM, Sun MZ, Zhao YX, Liang LY, Liu GJ, Zhang JH, Jia WS. Modulation of the root-sourced ABA signal along its way to the shoot in Vitis ripariaxVitis labrusca under water deficit. J Exp Bot, 2011, 62: 1731-1741[本文引用:1]
[53]
Carvalho D RA, FanourakisD, Correia MJ, Monteiro JA, Araújo-Alves J PL, Vasconcelos MW, Almeida D PF, HeuvelinkE, Carvalho S MP. Root-to-shoot ABA signaling does not contribute to genotypic variation in stomatal functioning induced by high relative air humidity. Environ Exp Bot, 2016, 123: 13-21[本文引用:2]
[54]
MiyawakiK, Matsumoto KitanoM, KakimotoT. Expression of cytokinin biosynthetic isopentenyltransferase genes in Arabidopsis: tissue specificity and regulation by auxin, cytokinin, and nitrate. Plant J, 2004, 37: 128-138[本文引用:1]
[55]
TakeiK, UedaN, AokiK, KuromoriT, HirayamaT, ShinozakiK, YamayaT, SakakibaraH. AtIPT3 is a key determinant of nitrate-dependent cytokinin biosynthesis in Arabidopsis. Plant Cell Physiol, 2004, 45: 1053-1062[本文引用:1]
[56]
NovákováM, MotykaV, Dobrev PI, MalbeckJ, GaudinováA, VankováR. Diurnal variation of cytokinin, auxin and abscisic acid levels in tobacco leaves. J Exp Bot, 2005, 56: 2877-2883[本文引用:1]
Xu ZJ, NakajimaM, SuzukiY, YamaguchiI. Cloning and characterization of the abscisic acid-specific glucosyltransferase gene from adzuki bean seedlings. Plant Physiol, 2002, 129: 1285-1295[本文引用:1]
[59]
McAdam S AM, Brodribb TJ. The evolution of mechanisms driving the stomatal response to vapor pressure deficit. Plant Physiol, 2015, 167: 833-843[本文引用:1]
[60]
NotaguchiM, OkamotoS. Dynamics of long-distance signaling via plant vascular tissues. Front Plant Sci, 2015, 6: 161[本文引用:1]
Lough TJ, Lucas WJ. Integrative plant biology: role of phloem long-distance macromolecular trafficking. Annu Rev Plant Biol, 2006, 57: 203-232[本文引用:1]
[66]
TurgeonR, WolfS. Phloem transport: cellular pathways and molecular trafficking. Annu Rev Plant Biol, 2009, 60: 207-221[本文引用:1]
[67]
Lucas WJ, GrooverA, LichtenbergerR, FurutaK, Yadav SR, HelariuttaY, He XQ, FukudaH, KangJ, Brady SM. The plant vascular system: evolution, development and functions. J Integr Plant Biol, 2013, 55: 294-388[本文引用:2]
[68]
Turnbull C GN, Lopez Cobollo RM. Heavy traffic in the fast lane: long-distance signalling by macromolecules. New Phytol, 2013, 198: 33-51[本文引用:1]
[69]
OkamotoS, SuzukiT, KawaguchiM, HigashiyamaT, MatsubayashiY. A comprehensive strategy for identifying long-distance mobile peptides in xylem sap. Plant J, 2015, 84: 611-620[本文引用:1]
 , 王逸茹, 安静, 王保民, 田晓莉
, 王逸茹, 安静, 王保民, 田晓莉





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}