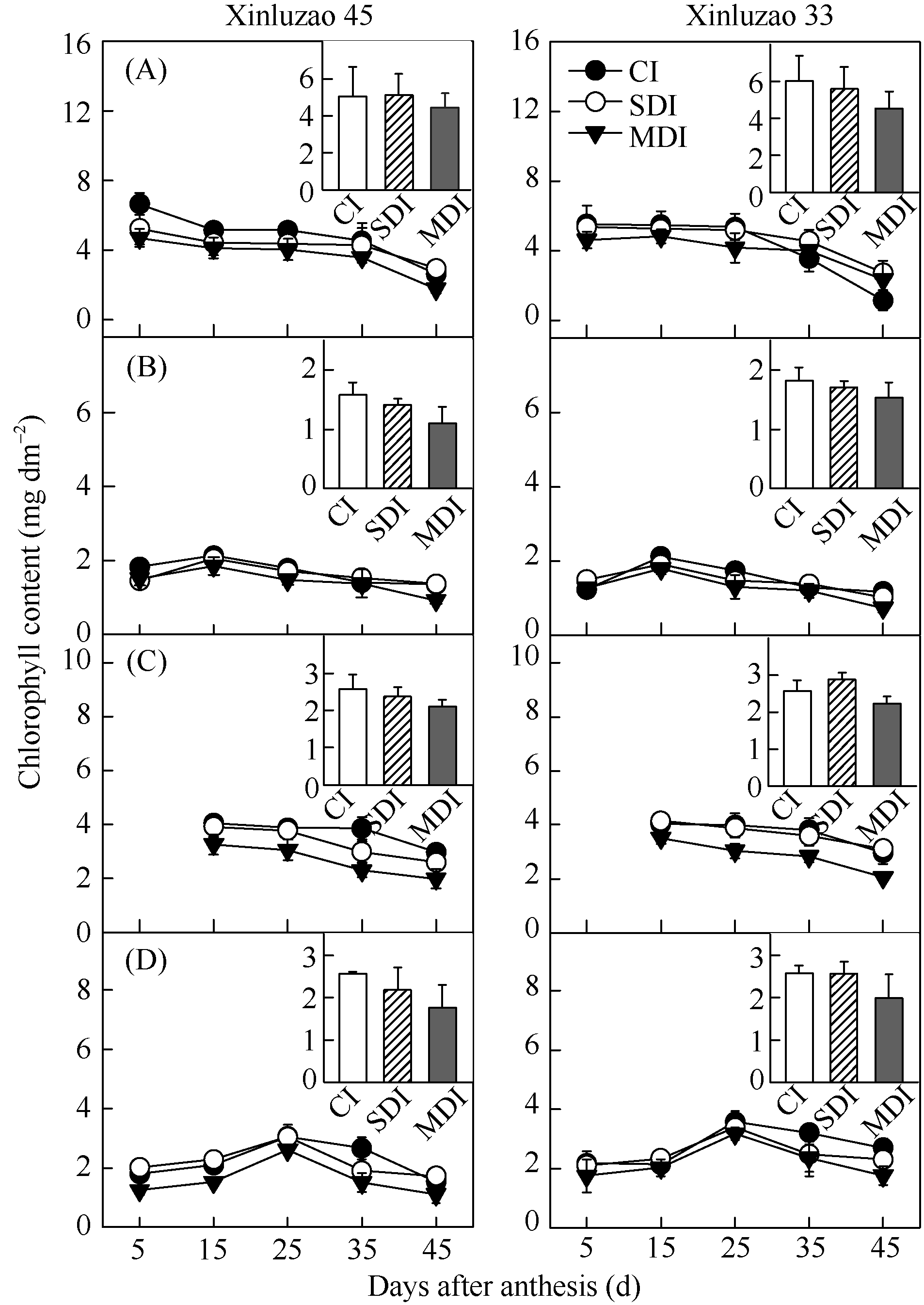
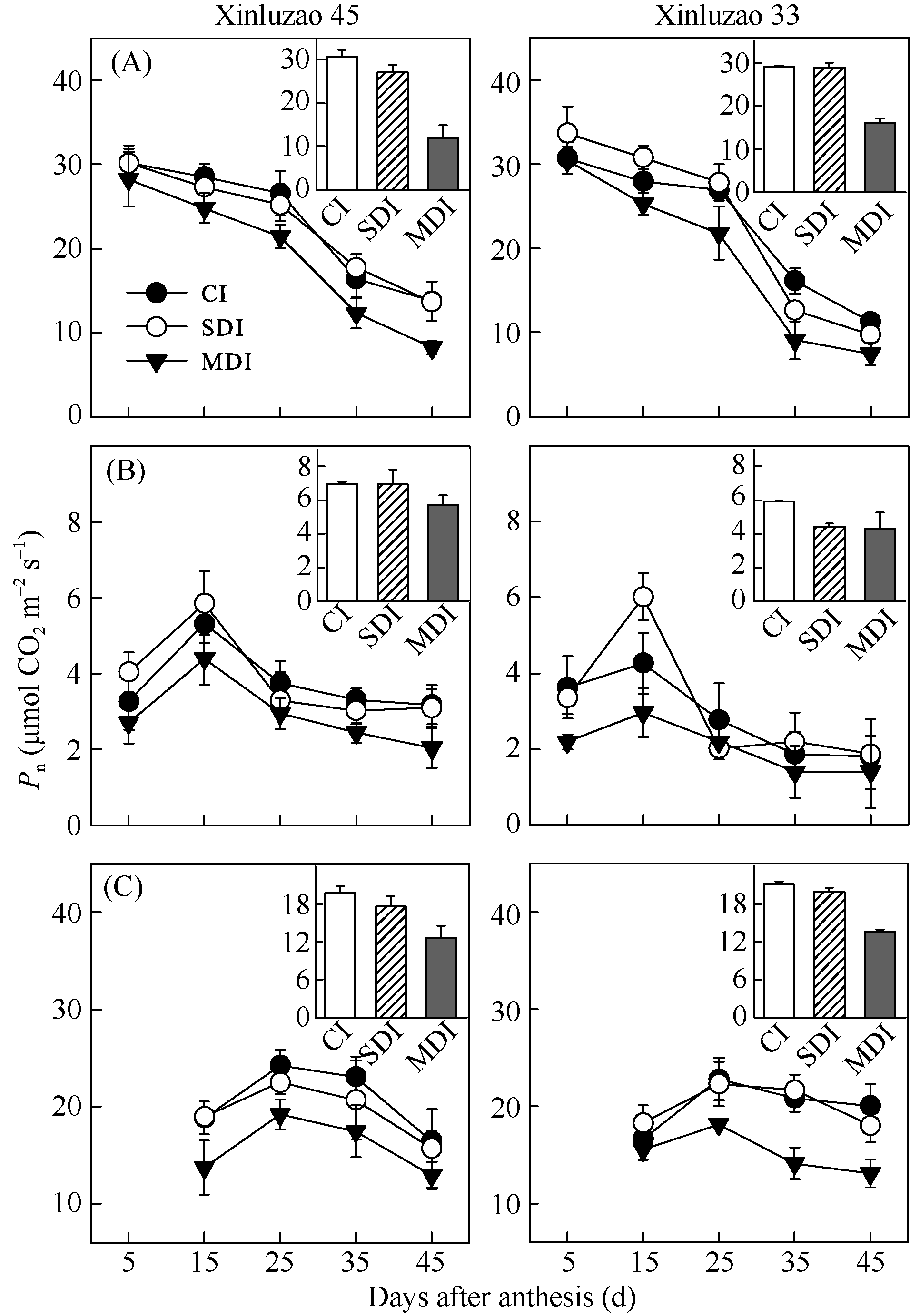
关键词:棉花; 非叶绿色器官; 光合能力; 相对贡献率; 水分亏缺 Photosynthetic Characteristics after Flowering and Contribution of Non-leaf Green Organs of Cotton to Yield under Mulching-drip Irrigation with Water Deficiency ZHAN Dong-Xia, ZHANG Chao, ZHANG Ya-Li, LUO Hong-Hai, GOU Ling, ZHANG Wang-Feng* Key Laboratory of Oasis Ecology Agriculture of Xinjiang Production and Construction Group / Agriculture College, Shihezi University, Shihezi 832003, China Fund:This study was supported by the National Natural Science Foundation of China (U1203283) and the Doctoral Foundation Program of Xinjiang Production and Construction Corps (2013BB002) AbstractLeaf is one of the main photosynthetic organs, while other green parts of plant also retain or develop chlorophyll and have photosynthesis. To better understand the whole plant photosynthesis production potential and contribution to cotton yield, we selected Xinluzao 33 and Xinluzao 45 (two common cultivars in Northern Xinjiang) with three irrigation treatments (CI, conventional irrigation; SDI, slight deficit irrigation; MDI, moderate deficit irrigation) to measure the chlorophyll content (Chl), net photosynthetic rate ( Pn), stomatal conductance ( gs) and photosynthate accumulation in leaf and non-leaf green organs during different growth stages and the contribution of non-leaf green organs to yield. The results showed that the Pn and Chl in non-leaf organs were relatively insensitive to soil moisture stress, decreasing by only a small amount between 25 and 45 days after anthesis. With reduction of water supply, the dry matter production in non-leaf green organs played more important roles in cotton yield formation. Cotton boll weight in the moderate deficit irrigation treatment decreased by 16.8% to 34.9% when the bolls (capsule walls plus bracts) were shaded and by 7.6% to 17.5% when the stalks were shaded. Hence, limiting-irrigation treatment is important to maintain high leaf photosynthetic rates. It is also important to develop the potential photosynthetic capacity of non-leaf green organs. This is especially important when leaf photosynthesis capacity declines due to leaf aging or water stress.
Keyword:Cotton; Non-leaf green organ; Photosynthesis; Relative contribution; Water deficiency Show Figures Show Figures
表1 不同滴灌量处理下棉花各绿色器官的气孔导度(2013) Table 1 Effect of water deficit on stomatal conductance (gs) in cotton organs during various times after anthesis in 2013 (mol m-2s-1)
花后时间 Days after anthesis
器官 Organ
新陆早45 号Xinluzao 45
新陆早33号 Xinluzao 33
CI
SDI
MDI
CI
SDI
MDI
5 d
叶片 Leaf
0.79± 0.02 a
0.68± 0.05 b
0.45± 0.03 c
0.76± 0.01 a
0.73± 0.08 a
0.46± 0.03 b
苞叶 Bract
0.09± 0.02 a
0.08± 0.02 b
0.06± 0.01 c
0.10± 0.02 a
0.09± 0.02 ab
0.07± 0.01 b
铃壳 Capsule wall
—
—
—
—
—
—
25 d
叶片 Leaf
0.49± 0.02 a
0.39± 0.09 b
0.14± 0.03 c
0.51± 0.00 a
0.48± 0.09 b
0.09± 0.03 c
苞叶 Bract
0.08± 0.01 a
0.06± 0.01 b
0.05± 0.01 c
0.09± 0.02 a
0.08± 0.02 a
0.06± 0.02 a
铃壳 Capsule wall
0.22± 0.02 a
0.18± 0.02 b
0.09± 0.01 c
0.26± 0.01 a
0.19± 0.04 b
0.10± 0.03 c
45 d
叶片 Leaf
0.19± 0.05 a
0.16± 0.03 b
0.13± 0.02 c
0.23± 0.05 a
0.18± 0.02 b
0.14± 0.03 c
苞叶 Bract
0.06± 0.02 a
0.04± 0.01 b
0.04± 0.01 b
0.05± 0.01 a
0.05± 0.01 a
0.04± 0.01 b
铃壳 Capsule wall
0.13± 0.03 a
0.09± 0.04 b
0.10± 0.03 b
0.18± 0.03 a
0.15± 0.04 b
0.07± 0.03 c
CI: conventional irrigation; SDI: slight deficit irrigation; MDI: moderate deficit irrigation. Comparing water treatments within a cultivar, values within a line followed by different letters are significantly different at the 0.05 probability level of probability according to LSD test. “ — ” means fruit was not sufficiently developed at that time so that the capsule wall could not be analyzed. CI: 常规滴灌量处理; SDI: 轻度水分亏缺处理; MDI: 中度水分亏缺处理。同一行同一品种不同小写字母表示各器官水分处理间在0.05水平上差异显著。“ — ” 表示铃太小无法测定。
表1 不同滴灌量处理下棉花各绿色器官的气孔导度(2013) Table 1 Effect of water deficit on stomatal conductance (gs) in cotton organs during various times after anthesis in 2013 (mol m-2s-1)
表2 不同滴灌量下棉花非叶绿色器官光合能力对铃重的相对贡献率 Table 2 Contribution of non-leaf organs to boll weight in cotton as affected by different water supplies (%)
品种 Cultivar
处理 Treatment
铃壳和苞叶 Capsule walls plus bracts
茎秆 Stalks
总贡献率 Total contribution
2012
新陆早33号 Xinluzao 33
CI
14.2± 1.0 b
9.2± 1.2 b
23.4± 2.7 c
SDI
14.8± 1.1 b
13.0± 2.9 a
27.8± 2.1 b
MDI
16.8± 1.4 a
14.3± 1.6 a
31.1± 3.3 a
新陆早45号 Xinluzao 45
CI
13.4± 1.3 b
9.6± 0.5 b
23.0± 1.8 b
SDI
26.5± 2.1 a
16.7± 1.8 a
43.2± 3.9 a
MDI
24.7± 1.9 a
17.5± 2.3 a
42.2± 2.4 a
2013
新陆早33号 Xinluzao 33
CI
18.9± 1.1 b
6.0± 0.5 b
18.6± 1.2 b
SDI
26.0± 1.8 a
10.7± 2.3 a
36.7± 2.7 a
MDI
27.4± 1.6 a
7.6± 1.5 b
35.0± 2.2 a
新陆早45号 Xinluzao 45
CI
29.7± 0.9 c
9.9± 0.8 b
39.6± 1.1 c
SDI
37.1± 2.0 a
15.4± 1.2 a
42.5± 2.6 b
MDI
34.9± 1.2 b
14.5± 1.1 a
49.4± 1.9 a
CI: conventional irrigation; SDI: slight deficit irrigation; MDI: moderate deficit irrigation. Comparing water treatments within a cultivar, values within a column followed by different letters are significantly different at the 0.05 probability level. CI: 常规滴灌量处理; SDI: 轻度水分亏缺处理; MDI: 中度水分亏缺处理。同一列相同品种不同小写字母表示在0.05水平上水分处理间差异显著。
表2 不同滴灌量下棉花非叶绿色器官光合能力对铃重的相对贡献率 Table 2 Contribution of non-leaf organs to boll weight in cotton as affected by different water supplies (%)
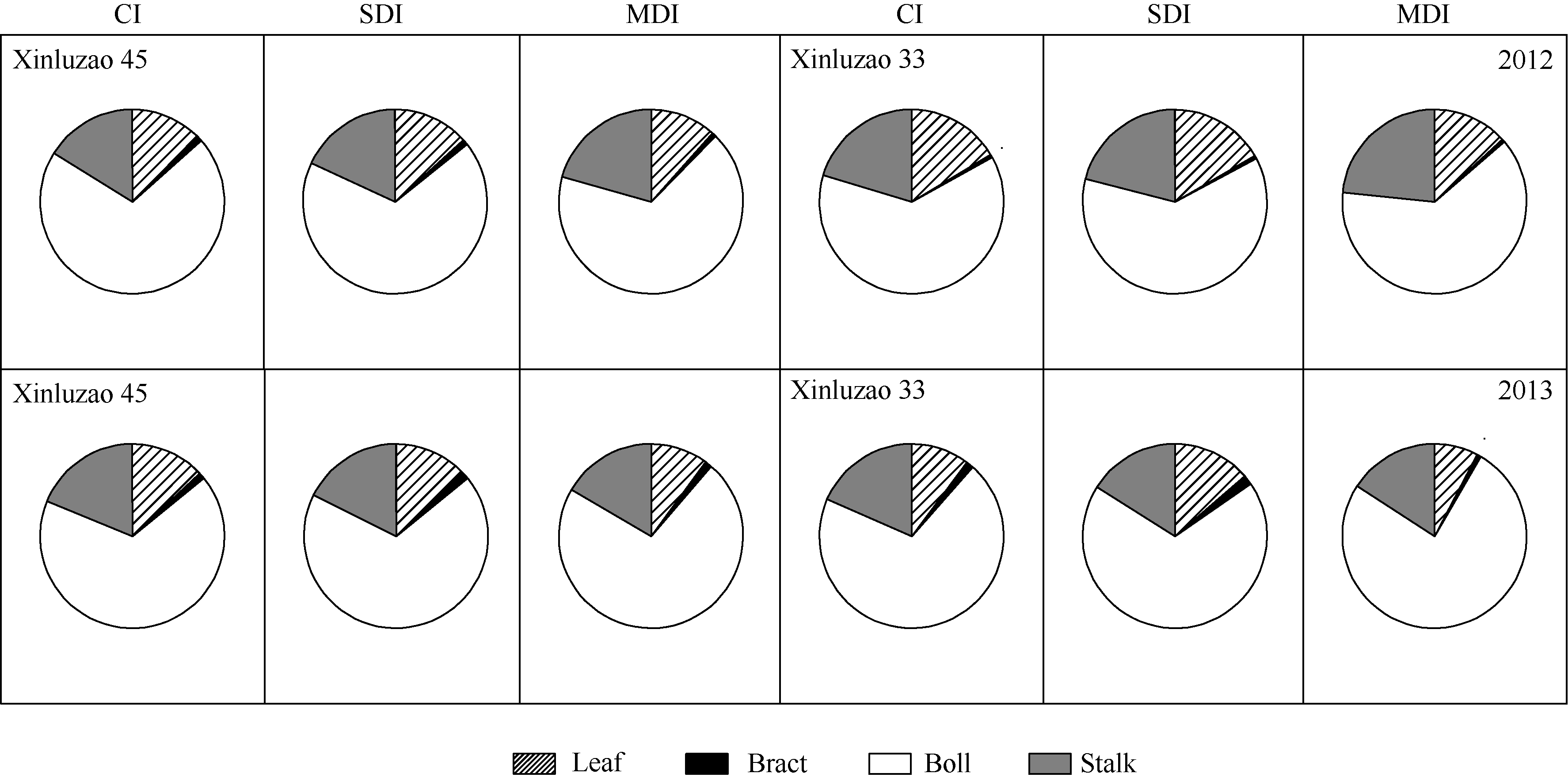
4 结论棉花生长后期, 随着水分亏缺程度的增加, 棉铃(铃壳和苞叶)和茎秆对铃重的相对贡献率增大。在生育后期叶片开始衰老时, 其光合能力迅速下降, 非叶绿色器官的光合作用对棉花产量的贡献增加, 尤其在水分亏缺条件下非叶绿色器官的光合功能期延长, 对棉铃的贡献增加。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
Araus JL, Brown HR, FebreroA, BortJ, Serret MD. Ear photosynthesis, carbon isotope discrimination and the contribution of respiratory CO2 to differences in grain mass in durum wheat. Plant Cell Environ, 1993, 16: 383-392[本文引用:4]
Zhang YP, Zhang YH, Wang ZM, Wang ZJ. Characteristics of canopy structure and contributions of non-leaf organs to yield in winter wheat under different irrigated conditions. Field Crops Res, 2011, 123: 187-195[本文引用:1]
[4]
Hu YY, Zhang YL, Luo HH, LiW, OguchiR, Fan DY, Chow WS, Zhang WF. Important photosynthetic contribution from the non-foliar green organs in cotton at the late growth stage. Planta, 2012, 235: 325-336[本文引用:4]
[5]
Evans LT, Rawson HM. Photosynthesis and respiration by the flag leaf and components of the ear during grain development in wheat. Aust J Biol Sci, 1970, 23: 245-254[本文引用:1]
[6]
RamH, SinghR. Chlorophyll content, photosynthetic rates and related enzyme activities in ear parts of two wheat cultivars differing in grain yield. Plant Physiol Biochem, 1982, 9: 94-102[本文引用:1]
[7]
Wang ZM, Wei AL, Zheng DM. Photosynthetic characteristics of non-leaf organs of winter wheat cultivars differing in ear type and their relationship with grain mass per ear. Photosynthetica, 2001, 39: 239-244[本文引用:1]
[8]
Salvador RJ, Pearce RB. Husk removal and its effects on maize grain yield. Crop Sci, 1988, 28: 961-964[本文引用:1]
[9]
ImaizumiN, UsudaH, NakamotoH, IshiharaK. Changes in the rate of photosynthesis during grain filling and the enzymatic activities associated with the photosynthetic carbon metabolism in rice panicles. Plant Cell Physiol, 1990, 31: 835-843[本文引用:1]
[10]
IshiharaK, KiyotaE, ImaizumiN. On the contribution of panicle photosynthesis to grain yield in rice plants. Jpn J Crop Sci, 1991, 60: 122-123[本文引用:1]
[11]
Crookston RK, O’Toole J, Ozbun J L. Characterization of the bean pod as a photosynthetic organ. Crop Sci, 1974, 14: 708-712[本文引用:1]
[12]
Xu HL, IshiiR. Wheat cultivar differences in photosynthetic response to low soil water potentials: 1. Maintenance of photosynthesis and leaf water potential. Jpn J Crop Sci, 1996, 65: 509-517[本文引用:1]
[13]
Martinez DE, Luquez VM, Bartoli CG, Guizmét JJ. Persistence of photosynthetic components and photochemical effect in ears of water-stressed wheat (Triticum aestivum). Physiol Plant, 2003, 119: 519-525[本文引用:1]
[14]
Tambussi EA, NoguésS, Araus JL. Ear of durum wheat under water stress water relations and photosynthesis metabolism. Planta, 2005, 221: 446-458[本文引用:1]
[15]
Tambussi EA, BortJ, Guiamet JJ, NoguesS, Araus JL. The photosynthetic role of ears in C3 cereals: metabolism, water use efficiency and contribution to grain yield. Plant Sci, 2007, 26: 1-16[本文引用:1]
[16]
Zhang YP, Zhang YH, Wang ZM, Wang ZJ. Characteristics of canopy structure and contributions of non-leaf organs to yield in winter wheat under different irrigated conditions. Field Crops Res, 2011, 123: 187-195[本文引用:1]
[17]
BortJ, FebreroA, AmaroT, Araus JL. Role of awns in ear water-use efficiency and grain weight in barley. Agronomie, 1994, 2: 133-139[本文引用:1]
[18]
AbbadH, Jaafrai SE, BortJ, Araus JL. Comparison of flag leaf and ear photosynthesis with biomass and grain yield of durum wheat under various water conditions and genotypes. Agronomie, 2004, 24: 19-28[本文引用:1]
[19]
魏爱丽, 王志敏, 陈斌, 翟志席, 张英华. 土壤干旱对小麦绿色器官光合电子传递和光合磷酸化活力的影响. 作物学报, 2004, 30: 487-490Wei AL, Wang ZM, ChenB, Zhai ZX, Zhang YH. Effect of soil drought on electron transport rate and photophosphorylation level of different green organs in wheat. Acta Agron Sin, 2004, 30: 487-490 (in Chinese with English abstract)[本文引用:1]
[20]
王志敏, 张英华, 张永平, 吴永成. 麦类作物穗器官的光合性能研究进展. 麦类作物学报, 2004, 24: 136-139Wang ZM, Zhang YH, Zhang YP, Wu YC. Review on photosynthetic performance of ear organs in Triticeae crops. J Triticeae Crops, 2004, 24: 136-139 (in Chinese with English abstract)[本文引用:1]
[21]
Wullschleger SD, Oosterhuis DM. Photosynthetic and respiratory activity of fruiting forms within the cotton canopy. Plant Physiol, 1990, 94: 463-469[本文引用:5]
[22]
Brown KJ. Translocation of carbohydrates in cotton: movement to the fruiting bodies. Ann Bot, 1968, 32: 703-713[本文引用:2]
[23]
Bondada BR, Oosterhuis DM. Comparative epidermal ultrastructure of cotton (Gossypium hirsutum L. ) leaf, bract and capsule wall. Ann Bot, 2000, 86: 1143-1152[本文引用:1]
[24]
Lichtenthaler HK. Chlorophylls and carotenoids: pigments of photosynthetic biomembranes. Methods Enzymol, 1987, 148: 350-382[本文引用:1]
[25]
Elmore CD. Contributions of the capsule wall and bracts to the developing cotton fruit. Crop Sci, 1973, 13: 751-752[本文引用:1]
[26]
Chapin FS. The mineral nutrition of wild plants. Annu Rev Ecol Syst, 1980, 11: 233-260[本文引用:1]
[27]
Bondada BR, Oosterhuis DM, Murphy JB, Kim KS. Effect of water stress on the epicuticular wax composition and ultrastructure of cotton leaf, bract, and boll. Environ Exp Bot, 1996, 36: 61-69[本文引用:1]
[28]
Van Iersel MW, Oosterhuis DM. Diurnal water relations of expand ing and full-sized cotton fruits and subtending leaves. Plant Cell Environ, 1995, 18: 807-812[本文引用:1]
[29]
Hu YY, Zhang YL, Yi XP, Zhan DX, Luo HH, Chow WS, Zhang WF. The relative contribution of non-foliar organs of cotton to yield and related physiological characteristics under water deficit. J Integr Agric, 2014, 13: 975-989[本文引用:2]
[30]
PooleI, Weyers J D B, Lawson T, Raven J A. Variations in stomatal density and index: implications for palaeoclimatic reconstructions. Plant Cell Environ, 1996, 19: 705-712[本文引用:1]
[31]
Zhan DX, YangY, Hu YY, Zhang YL, Luo HH, Zhang WF. Contributions of nonleaf organs to the yield of cotton grown with different water supply. Sci World J, 2014, DOI: DOI:10.1155/2014/602747[本文引用:1]
[32]
Hu YY, OguchiR, Yamori W, von Caemmerer S, Chow W S, Zhang W F. Cotton bracts are adapted to a micro-environment of concentrated CO2 produced by rapid fruit respiration. Ann Bot, 2013, 112: 31-40[本文引用:1]
[33]
胡立勇, 单文燕, 王维金. 油菜结实特性与库源关系的研究. 中国油料作物学报, 2002, 24(2): 37-42Hu LY, Shan WY, Wang WJ. Characteristics of seed set and source-sink relation of Brassica napus. Chin J Oil Crop Sci, 2002, 24(2): 37-42 (in Chinese with English abstract)[本文引用:1]
[34]
冷锁虎, 朱耕如, 邓秀兰. 油菜籽粒干物质来源的研究. 作物学报, 1992, 18: 250-257Leng SH, Zhu GR, Deng XL. Studies on the sources of the dry matter in the seed of rapeseed. Acta Agron Sin, 1992, 18: 250-257 (in Chinese with English abstract)[本文引用:1]
[35]
Mogensen VO, Jensen CR, MortensenG, Andersen MN, Schjoerring JK, Thage JH, KoribidisJ. Pod photosynthesis and drought adaptation of field grown rape (Brassica napus L. ). Eur J Agron, 1997, 6: 295-307[本文引用:1]
[36]
杜明伟, 冯国艺, 姚炎帝, 罗宏海, 张亚黎, 夏东利, 张旺锋. 杂交棉标杂A1和石杂2号超高产冠层特性及其与群体光合生产的关系. 作物学报, 2009, 35: 1068-1077Du MW, Feng GY, Yao YD, Luo HH, Zhang YL, Xia DL, Zhang WF. Canopy characteristics and its correlation with photosynthesis of super high-yielding hybrid cotton Biaoza AI and Shiza 2. Acta Agron Sin, 2009, 35: 1068-1077 (in Chinese with English abstract)[本文引用:1]
[37]
Biscoe PV, Scott RK, Monteith JL. Barley and its environment: III. Carbon budget of the stand . J Appl Ecol, 1975, 12: 269-270[本文引用:1]



{kind=link}
{kind=link}
{kind=link}