关键词:簇毛麦-小麦易位系; 转录组; 白粉病抗性; 分子标记 Developing of Specific Transcription Sequences P21461 and P33259 on Dasypyrum villosum 6VS and Application of Molecular Markers in Identifying Wheat- D. villosumBreeding Materials with Powdery Mildew Resistance LIU Chang, LI Shi-Jin, WANG Ke, YE Xing-Guo, LIN Zhi-Shan* National Key Facility for Crop Gene Resources and Genetic Improvement / Key Laboratory of Crop Genetics and Breeding, Ministry of Agriculture / Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China Fund:This study was supported by the National Research and Development Program (2016YFD0102002) and the Agricultural Science and Technology Innovation Program of CAAS. Abstract Dasypyrum villosum carries powdery mildew (PM) resistance gene Pm21 and PmV on its chromosome arms of 6V#2S and 6V#4S, respectively. The two resistant genes are co-segregated with exogenous chromosome arms in the offspring of hybrids between translocation lines and common wheat varieties. Developing polymorphic expression sequences to identify the two exogenous chromosomal arms is of great significance for their application in genetics and breeding, especially for 6V#4S chromosome, on which hereditary informations relatively lack. In this study, transcriptome of leaves of 6V#4S·6DL translocation line Pm97033 and common wheat line Wan7107 inoculated with Bgt was used as resources for sequences screening. As results, two unigenes of P21461 and P33259 from 6V#4S were identified by differential gene screening, synteny analysis, genomic DNA amplification and products sequencing of D. villosum accordingly. By using a pair of designed primer P461-5 based on P21461 sequence, it was found that there were 30 bp of InDel and 4 nt polymorphism between chromosomal arms of 6V#2S and 6V#4S. By the newly developed marker P461-5a, which retains the InDel polymorphism and truncates the product size amplified by P461-5, several wheat varieties and advanced lines with strong resistance to powdery mildew were identified to contain Pm21 or PmV. This marker showed potential application in discriminating D. villosum resources and assisted selection breeding for PM resistance in wheat. A marker P259-1 developed according to sequence of P33259 could specifically amplify a fragment from the wheat lines containing 6V#4S chromosome arm, but not amplify any fragment in the wheat lines containing 6V#2S·6AL translocation chromosome. Thereby, P259-1 can be used as a 6V#4S·6DL specific molecular marker in wheat backgrounds. The qRT-PCR assay showed that the expression of P21461 was not induced by PM. While transcriptional levels of P33259 increased about two times at 12 h and 24 h after inoculating the pathogens, suggesting that it might be involved in early interaction between Pm97033 and the pathogen.
Keyword: Dasypyrum villosum- Triticum aestivum translocation line; Transcriptome; Powdery mildew resistance; Molecular markers Show Figures Show Figures
S and R stand for sensitivity and resistance to powdery mildew, respectively. 92R178, 92R133, 88-128/Nannong P045, 92R137, and 92R179 carry 6V#2S· 6AL, while 93N40 carries 6V#4S· 6DL. Resistant advanced lines, provided by Prof. Xiao Chen from the Institute of Crop Science, Chinese Academy of Agricultural Science, are marked with “ ?” . S表示感病, R表示抗病。92R178、92R133、88-128/南农P045、92R137和92R179携带6V#2S· 6AL, 93N40携带6V#4S· 6DL。标?的为抗病高代品系, 由中国农业科学院作物科学研究所陈孝研究员提供。
表1 试验材料的名称、系谱/来源及对白粉病菌的反应 Table 1 Name, pedigree/origin, and reaction to powdery mildew of plant materials used in the study
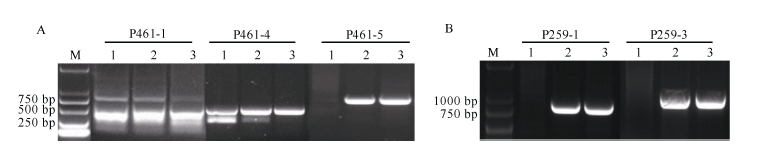
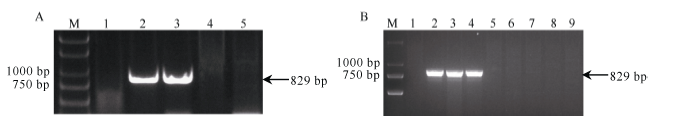
图2 引物P461-5在携带6V#2S和6V#4S的小麦易位系及已知系谱的抗白粉病小麦品种中的扩增谱带Fig. 2 Amplification profile of primer P461-5 in the 6V#2S and 6V#4S translocation lines and wheat varieties with known pedigree and powdery mildew resistance
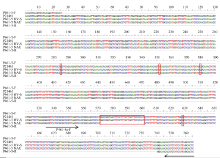
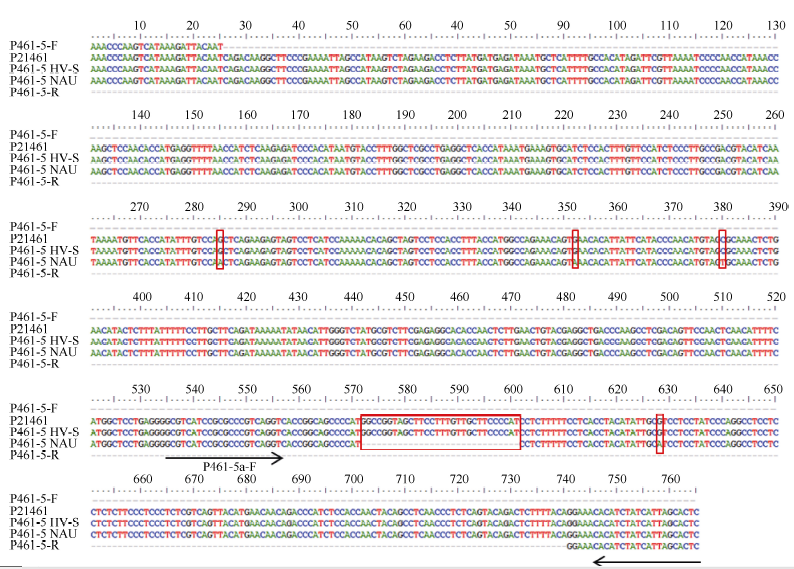
图3 引物P461-5在簇毛麦No.1026及6V#2S· 6AL易位系10SR3124中的扩增片段与原转录组序列的核苷酸序列比对图Fig. 3 Alignment of nucleotide sequences between the original transcriptome and the amplified fragments in D. villosum No.1026 and the 6V#2S· 6AL translocation line 10SR3124 using primer P461-5
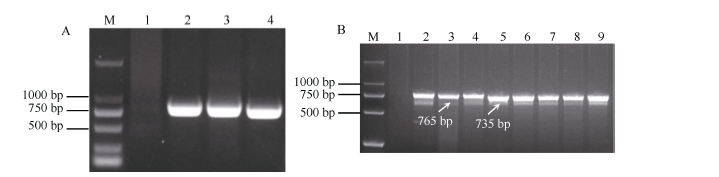
图4 引物P259-1在携带6V#2S和6V#4S的小麦易位系及已知系谱的抗白粉病小麦品种中的扩增谱带Fig. 4 Amplification profile of primer P259-1 in the 6V#2S and 6V#4S translocation lines and wheat varieties with known pedigree and powdery mildew resistance
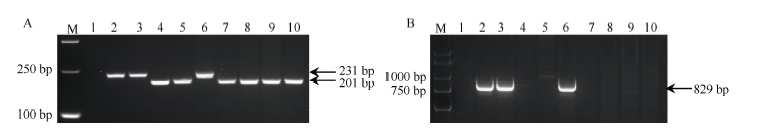
图5 引物P461-5a (A)和P259-1 (B)在不同小麦易位系、簇毛麦亲本及抗白粉病高代品系中的扩增Fig. 5 Amplification and identification of different wheat translocation lines, D. villosum parents, and wheat advanced lines with PM-resistance using primers P461-5a (A) and P259-1 (B)
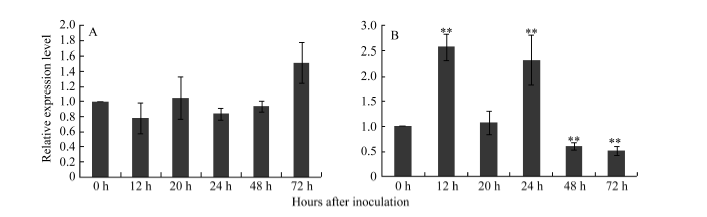
图6 Pm97033接种白粉病菌后72 h内P21461 (A)和P33259基因(B)的表达模式Fig. 6 Expression patterns of P21461 (A) and P33259 (B) in Pm97033 after inoculating Blumeria graminis f. sp. tritici (Bgt) for 72 hours
4 结论利用转录组测序数据成功筛选鉴定了2个来自Pm97033中簇毛麦6V#4S染色体臂的基因片段P21461和P33259, 将P21461转化成能在不同小麦遗传背景中区分6V#2S和6V#4S染色体臂的特异分子标记P461-5a, 而基于P33259的序列开发的标记P259-1可特异跟踪小麦遗传背景中的6V#4S· 6DL染色体, 并将2个标记用于鉴定育成的不同小麦-簇毛麦抗白粉病品种(系)。 The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
De PaceC, Qualset CO. Mating system and genetic differentiation in Dasypyrum villosum(Poaceae) in Italy. Plant Syst Evol, 1995, 197: 123-147[本文引用:1]
[2]
FrederiksenS. Taxonomic studies in Dasypyrum(Poaceae). Nord J Bot, 1991, 11: 135-142[本文引用:1]
[3]
GradzielewskaA. The genus Dasypyrum: part 2. Dasypyrum villosum—a wild species used in wheat improvement. Euphytica, 2006, 152: 441-454[本文引用:1]
[4]
Sears ER. Addition of the genome of Haynaldia villosa to Triticum aestivum. Am J Bot, 1953, 40: 168-174[本文引用:1]
[5]
LiuC, LiuW, WilsonJ, FriebeB, Gill BS. Development of a set of compensating Triticum aestivum- Dasypyrum villosum robertsonian translocation lines. Genome, 2011, 54: 836-844[本文引用:3]
[6]
Liu DJ, Chen PD, Pei GZ, Wang YL, Qiu BX, Wang SL. Transfer of Haynaldia villosa chromosomes into Triticum aestivum. In: Miller T E, Koebner R M D, eds. Proceeding of the 7th International Wheat Genetics Symposium. Cambridge, UK, 1988, pp355-361[本文引用:1]
[7]
Qi LL, Wang SL, Chen PD, Liu DJ, Gill BS. Identification and physical mapping of three Haynaldia villosa chromosome-6V deletion lines. Theor Appl Genet, 1998, 97: 1042-1046[本文引用:2]
[8]
Lin ZS, Zhang YL, Wang MJ, Li JR, WangK, ChenX, Xu QF, Zhang XS, Ye XG. Isolation and molecular analysis of genes Stpk-V2and Stpk-V3 homologous to powdery mildew resistance gene Stpk-V in a Dasypyrum villosum accession and its derivatives. J Appl Genet, 2013, 54: 417-426[本文引用:2]
[9]
CaoA, XingL, WangX, YangX, WangW, SunY, QianC, NiJ, ChenY, LiuD, WangX, ChenP. Serine/threonine kinase gene Stpk-V, a key member of powdery mildew resistance gene Pm21, confers powdery mildew resistance in wheat. Proc Natl Acad Sci USA, 2011, 108: 7727-7732[本文引用:1]
[10]
Zhang JC, Zheng HY, Li YW, Li HJ, LiuX, Qin HJ, Dong LL, Wang DW. Coexpression network analysis of the genes regulated by two types of resistance responses to powdery mildew in wheat. Sci Rep, 2016, 6: 23805[本文引用:1]
[11]
Li AL, Zhang RZ, PanL, Tang LC, Zhao GY, Zhu MZ, Chu JF, Sun XH, WeiB, Zhang XQ, Jia JZ, MaoL. Transcriptome analysis of H2O2-treated wheat seedlings reveals a H2O2-responsive fatty acid desaturase gene participating in powdery mildew resistance. PLoS One, 2011, 6: e28810[本文引用:1]
[12]
EraymanM, TurktasM, AkdoganG, GurkokT, InalB, IshakogluE, IlhanE, UnverT. Transcriptome analysis of wheat inoculated with Fusarium graminearum. Front Plant Sci, 2015, 6: 867[本文引用:1]
[13]
贾昌路, 张瑶, 朱玲, 张锐. 转录组测序技术在生物测序中的应用研究进展. 分子植物育种, 2015, 13: 2388-2394Jia CL, ZhangY, ZhuL, ZhangR. Application progress of transcriptome sequencing technology in biological sequencing. Mol Plant Breed, 2015, 13: 2388-2394 (in Chinese with English abstract)[本文引用:1]
[14]
ZhangH, Yang YZ, Wang CY, LiuM, LiH, FuY, Wang YJ, Nie YB, Liu XL, Ji WQ. Large-scale transcriptome comparison reveals distinct gene activations in wheat responding to stripe rust and powdery mildew. BMC Genomics, 2014, 15: 898[本文引用:1]
[15]
ZhangH, Hu WG, Hao JL, Lv SK, Wang CY, TongW, Wang YJ, Wang YZ, Liu XL, Ji WQ. Genome-wide identification and functional prediction of novel and fungi-responsive lincRNAs in Triticum aestivum. BMC Genomics, 2016, 17: 238[本文引用:1]
[16]
Hao YB, WangT, WangK, Wang XJ, Fu YP, Huang LL, Kang ZS. Transcriptome analysis provides insights into the mechanisms underlying wheat plant resistance to stripe rust at the adult plant stage. PLoS One, 2016, 11: e0150717[本文引用:1]
[17]
Li QQ, Niu ZB, Bao YG, Tian QJ, Wang HG, Kong LR, Feng DS. Transcriptome analysis of genes related to resistance against powdery mildew in wheat- Thinopyrum alien addition disomic line germplasm SN6306. Gene, 2016, 590: 5-17[本文引用:3]
[18]
陈孝, 徐惠君, 杜丽璞, 尚立民, 韩彬, 施爱农, 肖世和. 利用组织培养技术向普通小麦导入簇毛麦抗白粉病基因的研究. 中国农业科学, 1996, 29(5): 1-8ChenX, Xu HJ, Du LP, Shang LM, HanB, Shi AN, Xiao SH. Transfer of gene resistant to powdery mildew from H. villosa to common wheat by tissue culture. Sci Agric Sin, 1996, 29(5): 1-8 (in Chinese with English abstract)[本文引用:1]
[19]
LiH, ChenX, Xin ZY, Ma YZ, Xu HJ, Chen XY, JiaX. Development and identification of wheat- Haynaldia villosa T6DL·6VS chromosome translocation lines conferring resistance to powdery mildew. Plant Breed, 2005, 124: 203-205[本文引用:1]
[20]
庄巧生. 中国小麦品种改良及系谱分析. 北京: 中国农业出版社, 2003. p 117Zhuang QS. Chinese Wheat Improvement and Pedigree Analysis. Beijing: China Agriculture Press, 2003. p 117 (in Chinese)[本文引用:1]
[21]
吴宏亚, 张伯桥, 汪尊杰, 程顺和. 优质弱筋抗白粉病小麦新品种扬麦22的选育及配套栽培技术. 江苏农业科学, 2013, 41: 109-112Wu HY, Zhang BQ, Wang ZJ, Cheng SH. Breeding and cultivation techniques of a new high quality of weak gluten wheat varieties Yangmai 22 resistance to powdery mildew. Jiangsu Agric Sci, 2013, 41: 109-112 (in Chinese)[本文引用:1]
[22]
Chen PD, Qi LL, ZhouB, Zhang SZ, Liu DJ. Development and molecular cytogenetic analysis of wheat- Haynaldia villosa 6VS/6AL translocation lines specifying resistance to powdery mildew. Theor Appl Genet, 1995, 91: 1125-1128[本文引用:1]
[23]
曾小莉, 黄耀跃, 王相权, 王仕林, 康建. 国审品种内麦836小麦栽培技术规程. 种子世界, 2015, (5): 51-52Zeng XL, Huang YY, Wang XQ, Wang SL, KangJ. Cultivation techniques of wheat about state approved varieties of Neimai 836. Seed World, 2015, (5): 51-52 (in Chinese)[本文引用:1]
张云龙, 王美蛟, 张悦, 褚翠萍, 林志珊, 徐琼芳, 叶兴国, 陈孝, 张宪省. 不同簇毛麦6VS染色体臂的白粉病抗性特异功能标记的开发及应用. 作物学报, 2012, 38: 1827-1832Zhang YL, Wang MJ, ZhangY, Chu CP, Lin ZS, Xu QF, Ye XG, ChenX, Zhang XS. Development and application of functional markers specific to powdery mildew resistance on chromosome arm 6VS from different origins of Haynaldia villosa. Acta Agron Sin, 2012, 38: 1827-1832 (in Chinese with English abstract)[本文引用:1]
[26]
陈艳玲, 孙凯. 小麦品种金禾9123高产稳产栽培管理措施. 中国种业, 2011, (5): 70-71Chen YL, SunK. High and stable yield cultivation and management measures of wheat variety Jinhe 9123. China Seed Ind, 2011, (5): 70-71 (in Chinese)[本文引用:1]
[27]
Paolacci AR, Tanzarella OA, PorcedduE, CiaffiM. Identification and validation of reference genes for quantitative RT-PCR normalization in wheat. BMC Mol Biol, 2009, 10: 11[本文引用:1]
[28]
Livak KJ, Schmittgen TD. Analysis of relative gene expression data using real-time quantitative PCR and the 2-ΔΔCt method. Methods, 2001, 25: 402-408[本文引用:1]
[29]
Bie TD, Zhao RH, Zhu SY, Chen SL, CenB, Zhang BQ, Gao DR, Jiang ZN, Chen TT, WangL, Wu RL, He HG. Development and characterization of marker MBH1simultaneously tagging genes Pm21and PmV conferring resistance to powdery mildew in wheat. Mol Breed, 2015, 35: 1-8[本文引用:1]
[30]
李桂萍, 陈佩度, 张守忠, 赵和. 小麦-簇毛麦6VS/6AL易位染色体对小麦农艺性状的影响. 植物遗传资源学报, 2011, 12: 744-749Li GP, Chen PD, Zhang SZ, ZhaoH. Effects of the 6VS/6AL translocation chromosome on agronomic characteristics of wheat. J Plant Genet Resour, 2011, 12: 744-749 (in Chinese with English abstract)[本文引用:1]
[31]
LiuC, Ye XG, Wang MJ, Li SJ, Lin ZS. Genetic behavior of Triticum aestivum-Dasypyrum villosum translocation chromosomes T6V#4S·6DL and T6V#2S·6AL carrying powdery mildew resistance. J Integr Agric, 2017, DOI: DOI:10.1016/S2095-3119(16)61568-X[本文引用:1]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}