网络出版日期:2017-07-01
摘要
关键词:玉米; 热激转录因子基因 ZmHsf25; 表达; 亚细胞定位; 耐热性
Cloning, Characteristics and Regulating Role in Thermotolerance of Heat Shock Transcription Factor ( ZmHsf25) in Zea maysL
ZHAO Li-Na
Abstract
Keyword:Maize; ZmHsf25; Expression; Subcelullar-localization; Thermotolerance
Show Figures
Show Figures







许多研究已表明, 植物热激蛋白(Hsp, heat shock protein)与植株耐热性的提高密切相关, 而植物热激转录因子(Hsf, heat shock transcription factor)对下游热激蛋白基因及耐热性相关基因的表达起关键调控作用, 是植物对高温胁迫反应的关键因子。自20世纪80年代后期首次克隆酵母Hsf以来, 相继获得动植物多个物种的Hsf基因[1, 2, 3]。热激转录因子在植物中普遍存在, 首个植物Hsf基因来自番茄[4]。与其他生物体相比, 植物中的Hsf基因是一个多基因家族, 依据结构分为A、B、C三族, 细分为多个亚族。Hsf家族基因数目因作物品种不同差异较大, 果蝇和酵母中只有1个, 而小麦中至少有56个[5], 从数量上远远多于已知的其他生物体。相比于其他植物, 对模式植物番茄和拟南芥Hsf基因功能和调控机制研究得比较深入, 研究对象主要集中在A1和A2亚族[6, 7, 8]。
关于B族Hsf的研究报道不多, 该族基因因不含激活结构域而不能直接激活下游基因的表达, 早期研究认为其作用更多表现为抑制[9, 10]。近年来, 越来越多的功能被逐渐报道。拟南芥中有5个B族Hsf, 其中HsfB2a和HsfB2b属于热诱导型, 主要在热激反应后期起作用, 但其热激诱导表达的前提是与HsfA1a和HsfA1b结合。HsfB1和HsfB2b定位在细胞核, 正常生长和热恢复阶段直接抑制热激转录因子HsfA2的活性, 进而抑制一系列Hsp基因的表达, 从而钝化热激反应。但在热激条件下, HsfB1和HsfB2b可能通过抑制某些干扰HsfA1转位入核的Hsp基因的表达, 从而促进HsfA1入核而激活HsfA1, 进而提高获得耐热性能力[10], 表现出正向调控功能。转鹰嘴豆CarHsfB2的拟南芥表现较强的抗旱和耐热性[11]。HsfB1和HsfB2b还参与pdf1.2的表达和抗病反应[12], 核中过表达HsfB1可导致细胞死亡[13]。HsfB2b组成型表达可降低节律相关基因Pseudo response regulator 7 (PRR7)的表达, 使得植株延迟开花, 下胚轴伸长, 因此在调控逆境条件下植株生物钟变化方面起重要作用[14]。番茄HsfB1作为共激活因子可增强HsfA1a和/或HsfA2的活性, 热激条件下可保持和恢复某些重要基因或持家基因的表达[15, 16]。另外, 过表达拟南芥HsfB4可特异性诱导根系发育[17], 水稻HsfB4b可通过结合特异HSE而调控病程防御反应[18]。可见, B族Hsf在植物抗逆、抗病过程中也发挥着重要的调控作用, 随着研究的逐步深入, 越来越多B族Hsf的特性和功能将得以解析。
通过生物信息学推测, 玉米中至少有30个Hsf成员, B族有7个, 其中4个基因同时含有核定位信号和胞质定位信号[3]。这些B族Hsf有什么样的特性和功能, 目前还没有相关报道。本实验室在研究玉米A1亚族基因的同时, 克隆了B族成员ZmHsf25, 拟初步分析其结构、表达和亚细胞定位特性, 及其耐热性调控功能, 以期为全面深入研究玉米热激转录因子家族基因提供理论基础和技术支撑。
1 材料与方法1.1 材料培养供试材料为玉米(Zea maysL.)自交系H21 (实验室备)。精选种子, 经表面消毒后, 以自来水反复冲洗, 浸泡吸胀12 h后, 播至Hoagland营养液中, 于28℃培养箱中暗培养。待玉米长到二叶一心时, 进行不同处理, 取样进行定量表达分析。
于2016年5月上旬田间穴播试材, 常规种植, 于开花盛期取功能叶、雌穗和花粉, 授粉2周后取幼胚, 以液氮速冻, 用于组织器官中基因定量分析。
1.2 胁迫处理将生长一致的幼苗分为5组, 分别将幼苗根部浸入溶液, 进行如下处理, 参照李慧聪等[19]的方法略加修改。(1)浸入42℃培养箱中预热的Hoagland营养液进行热胁迫处理(HS), 处理时间分别为10、20、30、40、50、60和240 min; (2)浸入含0.8 mmol L-1水杨酸(SA)的Hoagland溶液, 处理10、30、60、90、120和240 min; (3)浸入含10 mmol L-1 H2O2的Hoagland溶液, 处理时间同水杨酸; (4)浸入含0.8 mmol L-1水杨酸(SA)的Hoagland溶液预处理1.5 h, 然后移入新的含有SA的Hoagland溶液于42℃热处理50 min; (5)浸入含10 mmol L-1 H2O2的Hoagland溶液预处理1.5 h, 然后移入新的含有H2O2的Hoagland溶液于42℃热处理50 min。取样并以液氮速冻, 同时取28℃下正常生长的幼苗作对照。
1.3 基因克隆与测序采用上海华舜生物工程公司生产的RNArose Reagent Systems试剂盒提取总RNA, 经DNaseI (TaKaRa, 大连)处理除去残留的基因组DNA, 然后取1 μ g合成cDNA第1链(反转录试剂盒, Invitrogen, 美国)。依据玉米B73基因组信息设计特异引物(正向引物5° -TCCGTTGCCTGATGGCGTTGCC-3° ; 反向引物5° -TCATTTCAGCCCTAGACTGAGGC-3° ), 利用高保真酶pyrobest (TaKaRa, 大连)进行PCR扩增。PCR反应体系(25 µ L)含10× pyrobest buffer 2.5 µ L、dNTP mixture (2.5 mmol L-1) 2 µ L、1st strand cDNA 2 µ L、20 µ mol L-1 正向引物0.25 µ L、20 µ mol L-1 反向引物 0.25 µ L、Pyrobest DNA polymerase 0.25 µ L和ddH2O 17.75 µ L。反应程序为98℃ 10 s, 55℃ 15 s, 72℃ 2 min, 30个循环。将扩增产物连接T载体后(pEasy-blunt simple cloning kit, TransGen Biotech, 北京), 送上海生工生物工程技术服务有限公司测序。
1.4 qRT-PCR分析依据Lin等[20]对玉米Hsf的分析合成ZmHsf25的特异引物(正向引物5'-AGTGAAGCCAATGCTG TTCG-3'; 反向引物5° -CTTGTTCCTCGTCGTTGTC C-3° ), 和玉米内参基因β -actin的特异性引物(正向引物5° -TGTCCATCACTTGTGAAGCCTCCT-3° ; 反向引物5° -ACGACCTTAGCCAATATCGCACCA-3° )。PCR体系(20 µ L)含: SYBR Premix Ex TaqII 10 µ L、10 µ mol L-1 正向引物 0.8 µ L、10 µ mol L-1反向引物 0.8 µ L、1st strand cDNA 1 µ L和ddH2O 7.4 µ L。反应在7500 Real-time PCR System (Applied Biosystems, USA)上进行, 反应程序: 预变性95℃ 10 min, 变性 95℃ 5 s, 退火/延伸60℃ 1 min, 45个循环。反应结束后, 采用2-Δ Δ Ct分析数据。每组试验设3个生物学样本, 每个生物学样本设3次技术重复。数据为3个生物学重复平均值± 标准误, 组织表达试验以幼根表达量为1, 其余设置0 min的表达量为1。
1.5 ZmHsf25亚细胞定位观察带有绿色荧光蛋白(GFP)基因的植物表达载体pJIT163-hGFP (中国科学院遗传与发育生物学研究所范仁春博士提供)通过35S启动子驱动目的基因和GFP基因的融合表达[21], 显示目的蛋白的亚细胞定位。扩增ZmHsf25编码区, 产物经限制性内切酶Hind III和BamH I消化后, 连接到表达载体pJIT163-hGFP上。参照李慧聪等[22]方法进行金粉包埋和基因枪转化。经转化的洋葱表皮细胞于22℃暗培养16 h, 然后放入浓度为10 µ g mL-1的4’ , 6-二脒基-2-苯基吲哚(4’ , 6-diamidino-2-phenylindole, DAPI)染色液中染色3~5 min, 用生理盐水冲洗干净, 于激光共聚焦显微镜(Zeiss META510)下观察荧光。
1.6 酵母表达载体构建酵母表达载体pYES2 (Invitrogen, 美国)用于酿酒酵母中目的蛋白的表达, 其特点在于GAL1启动子能够在酿酒酵母中被半乳糖高水平诱导从而驱动目的蛋白表达, 同时能够被葡萄糖抑制表达, 可利用URA3基因筛选带有ura3基因型的酵母宿主菌株转化子。结合ClonExpress II重组反应系统(诺唯赞生物科技有限公司)设计扩增ZmHsf25特异引物(ZmHsf25-F: 5° -GGGAATATTAAGCTTGGTACCAT GGCGTTGCCGGCGGCG-3° ; ZmHsf25-R: 5° -TGAT GGATATCTGCAGAATTCTCATTTCAGCCCTAGACT-3° ), 利用高保真PCR聚合酶进行扩增, 获得ZmHsf25的PCR产物。利用限制性内切酶Kpn I和EcoR I (NEB)对载体pYES2进行酶切, 电泳后回收得到线性化载体。将PCR产物与线性化载体按1∶ 2的摩尔比混合, 利用ClonExpress II快速克隆技术进行重组反应。于冰水浴中配制如下反应体系即4 µ L 5× ClonExpress II buffer, 50~200 ng线性化载体, 20~200 ng插入片段扩增产物, 2 µ L Exnase II, 加无菌水至总体积2 µ L。混匀各组分后, 于37℃反应30 min, 立即置冰浴中冷却5 min, 然后利用热激法将反应产物直接转化大肠杆菌TOP10感受态细胞, 37℃倒置培养过夜。用无菌的牙签将单个菌落挑至100 µ L新鲜的LB培养基中, 混匀, 取2 µ L作为模板进行PCR扩增, 根据电泳条带大小选择正确的克隆进行序列测定。
1.7 ZmHsf25在酵母中转化及耐热性鉴定参照Gietz等[23]的方法, 将测序正确的重组载体转化酵母INVSc1感受态细胞, 然后将细胞均匀涂抹在SC-Glu-Ura-筛选平板上, 30℃培养2~3 d。采用菌落PCR方法鉴定阳性克隆。
分别将重组载体ZmHsf25-pYES2和空载体pYES2转化酵母细胞INVSc1, 筛选阳性酵母克隆。分别挑选2个菌系的阳性克隆, 以SC-Glu-Ura-液体培养基振荡(250转 min-1)培养过夜, 检测OD600值。用SC-Glu-Ura-液体培养基将酵母菌液稀释到0.2, 重新振荡(200转 min-1)培养2~3 h, OD600值达到0.4~0.8之间即酵母指数生长期, 离心收集菌体, 无菌水洗涤2遍, 除去培养基, 然后梯度稀释到0.10、0.05和0.01共3个OD值, 分成2组, 一组作为对照, 另一组用50℃水浴热激处理45 min。吸取8 μ L, 分别点在加入半乳糖的SC-Gal-Ura-固体培养基上, 30℃生长2~3 d, 观察并照相。另外, 将指数生长期的酵母细胞稀释到SC-Gal-Ura-液体培养基中, 分成2组, 一组作为对照, 另一组用50℃水浴中热激处理45 min, 振荡(200转 min-1)培养24 h, 期间每间隔3 h测定一次OD600值, 计算生长速率。采用Microsoft Excel分析数据, 计算标准误差。采用SPSS统计方法分析差异显著性。
2 结果与分析2.1 玉米热激转录因子基因ZmHsf25的克隆与结构分析 将二叶一心玉米幼苗于42℃热胁迫处理1 h, 提取叶片总RNA, 以此为模板, 利用特异性引物, 扩增获得ZmHsf25基因的完整编码序列(图1), 序列全长957 bp, 编码318个氨基酸残基。ZmHsf25蛋白序列包含DNA结合结构域(DNA binding domain, DBD, NCBI网站分析, 下画线部分)、核定位信号序列(nuclear localization signal, NLS, NucPred软件分析)和核输出信号(nuclear export signal, NES, NetNES软件分析)序列。DNA结合结构域是Hsf的典型结构特征; 根据核定位信号序列推测ZmHsf25被定位于细胞核; 核输出信号序列的存在表明在某些情况下ZmHsf25可能会穿梭出核发挥功能。利用DNAMAN软件分析表明, ZmHsf25蛋白与高粱Sb060g025710 (序列号为XP_002446950.1)、谷子SiHSFB-2a-like (序列号为XP_004976526.1)和水稻OSIGBa0103M18.7 (序列号为CAH66855.1)的相似性分别为92%、81%和77% (图2)。
图1
Fig. 1
| Figure OptionViewDownloadNew Window | |
 | 图1 玉米ZmHsf25的核苷酸和氨基酸序列(下画线部分: Hsf家族保守DNA结合域; NLS: 核定位信号序列; NES: 核输出信号序列。)Fig. 1 Nucleotide and amino acid sequences of ZmHsf25 from maize(Underline: the conserved DNA binding domain of Hsf family; NLS: nuclear localization signal; NES: nuclear export signal.) |
图2
Fig. 2
| Figure OptionViewDownloadNew Window | |
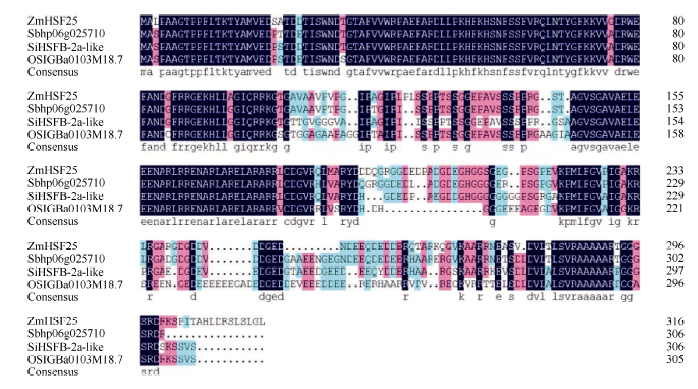 | 图2 玉米ZmHsf25蛋白与其他蛋白的相似性比较Fig. 2 Alignment of the putative amino acid encoded by ZmHsf25 and other proteins from different species |
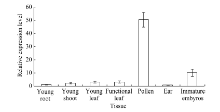
2.2 ZmHsf25基因在玉米不同组织器官中的表达分析通过分析玉米幼苗期和开花盛期不同组织器官中ZmHsf25的定量表达发现, 正常生长条件下, ZmHsf25在玉米根系中表达最低, 幼嫩花粉中表达量最高, 显著高于其他器官, 达根系的50倍, 其次是幼胚, 约为根系表达量的10倍(图3)。
图3
Fig. 3
| Figure OptionViewDownloadNew Window | |
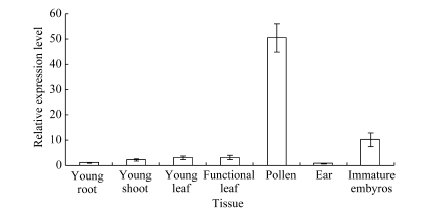 | 图3 正常生长条件下幼苗期和开花期不同组织器官中ZmHsf25的相对表达水平(每个样本设3次重复, 数据为3个生物学重复平均值± 标准误, 组织表达试验以幼根表达量为1。)Fig. 3 Expression levels of ZmHsf25 in different tissues and organs at seedling and anthesis stages under normal growth conditions(There are three replicates for each sample and the data are mean± standard error. The expression level 1 is the expression level in roots.) |
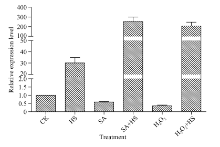
2.3 不同逆境胁迫对ZmHsf25表达的影响 42℃热胁迫能显著上调玉米幼苗叶片(图4-A)和根系(图4-B) ZmHsf25的表达, 处理50 min时达到峰值, 之后逐渐降低, 叶片最大值超过根系的2倍。正常生长条件下, 0.8 mmol L-1 SA处理下调玉米根系ZmHsf25的表达(图5-B), 对叶片基因表达影响不大(图5-A); 10 mmol L-1 H2O2处理对根系(图6-B)和叶片(图6-A)基因表达均表现为下调, 根系在240 min时恢复至初始值; 如果用0.8 mmol L-1 SA或10 mmol L-1 H2O2分别预处理幼苗1.5 h, 然后热激50 min, 叶片ZmHsf25的表达被大幅度上调(图7)。表明在热激条件下ZmHsf25可能参与SA和H2O2介导的信号通路。
图4
Fig. 4
| Figure OptionViewDownloadNew Window | |
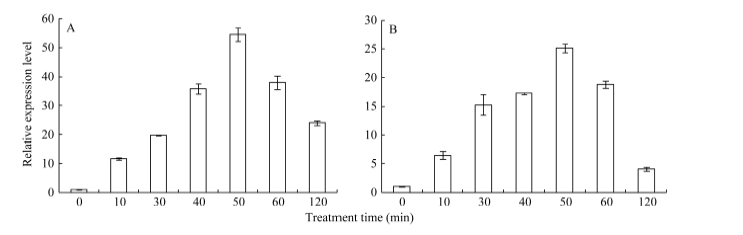 | 图4 42° C热处理对玉米ZmHsf25表达的影响(A:ZmHsf25在玉米叶片中的表达; B: ZmHsf25在玉米根系中的表达。)Fig. 4 Expression levels ofZmHsf25 in leaves and roots of the maize seedlings subjected to heat shock at 42° C(A: ZmHsf25expression levels in maize leaves; B: ZmHsf25expression levels in maize roots.) |
图5
Fig. 5
| Figure OptionViewDownloadNew Window | |
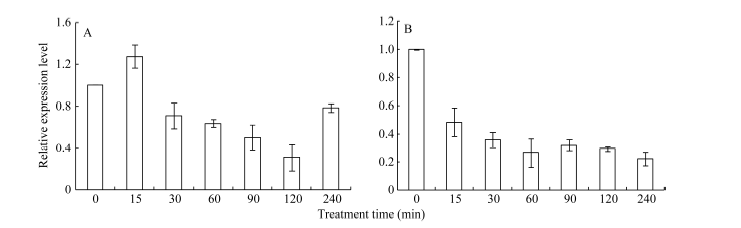 | 图5 0.8 mmol L-1水杨酸处理对玉米幼苗ZmHsf25表达的影响(A:ZmHsf25在玉米叶片中的表达; B: ZmHsf25在玉米根系中的表达。)Fig. 5 Expression levels of ZmHsf25 in leaves and roots of maize seedlings treated with 0.8 mmol L-1 SA(A: ZmHsf25expression levels in maize leaves; B: ZmHsf25expression levels in maize roots.) |
图6
Fig. 6
| Figure OptionViewDownloadNew Window | |
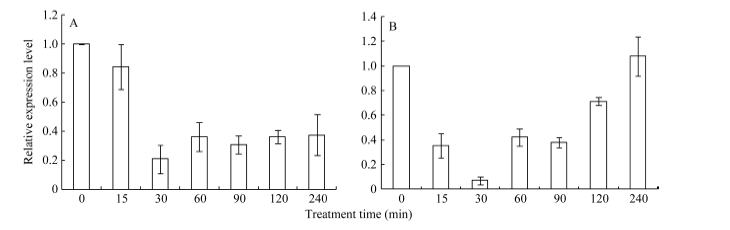 | 图6 10 mmol L-1H2O2处理对玉米幼苗ZmHsf25表达的影响(A:ZmHsf25在玉米叶片中的表达; B: ZmHsf25在玉米根系中的表达。)Fig. 6 Expression levels of ZmHsf25 in leaves and roots of the maize seedlings treated with 10 mmol L-1 H2O2(A: ZmHsf25expression levels in maize leaves; B: ZmHsf25expression levels in maize roots.) |
图7
Fig. 7
| Figure OptionViewDownloadNew Window | |
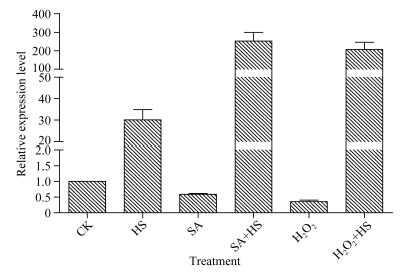 | 图7 热激条件下水杨酸和H2O2预处理对玉米叶片ZmHsf25表达的影响Fig. 7 Expression levels of ZmHsf25 in leaves of maize seedlings under heat shock at 42° C pretreated with 0.8 mmol L-1 SA and 10 mmol L-1 H2O2, respectively |
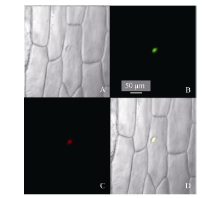
2.4 玉米ZmHsf25蛋白的亚细胞定位构建瞬时表达载体pJIT163-ZmHsf25-hGFP并转化洋葱内表皮, 通过融合基因表达显示目的蛋白的亚细胞定位。利用激光共聚焦显微镜观察荧光发现, 正常条件下, 观察到GFP绿色荧光只分布在细胞核, 且与核特异染料DAPI荧光完全重合(图8)。表明ZmHsf25蛋白定位于细胞核。
2.5 玉米ZmHsf25在酵母中耐热性鉴定 通过将玉米ZmHsf25在酵母中转化并进行热激处理, 观察酵母生长发现, 在正常生长条件下, 在半乳糖诱导固体培养基上, 转化ZmHsf25与转化空载体pYES2的酵母细胞的长势无明显差异(图9-A); 经50℃热激处理45 min后, 转基因酵母细胞和转化空载体对照的生长速度均受到抑制, 但是前者的受抑制程度明显小于后者(图9-B)。酵母细胞生长速率实验结果也充分证明了上述观察结果(图10-A, B)。这表明, ZmHsf25能在酵母细胞中被诱导表达, 且能显著提高酵母细胞的耐热性。
图8
Fig. 8
| Figure OptionViewDownloadNew Window | |
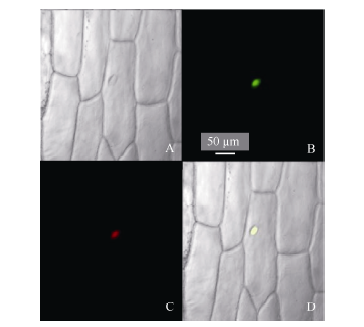 | 图8 玉米ZmHsf25蛋白质在洋葱表皮细胞中的亚细胞定位(A: 明场; B: GFP绿色荧光; C: 细胞核DAPI红色荧光; D: 叠加图像。)Fig. 8 Subcellular localization of ZmHsf25 in onion epidermal cells under normal conditions(A: bright field; B: green fluorescence of GFP; C: red fluorescence of DAPI; D: merged images.) |
图9
Fig. 9
| Figure OptionViewDownloadNew Window | |
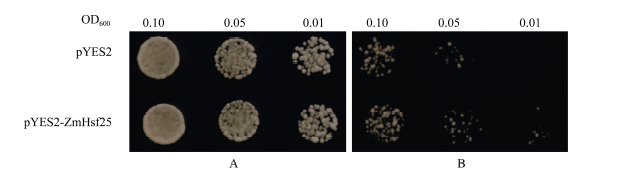 | 图9 50℃热激处理后转基因酵母耐热性观察(A: 正常培养; B: 50° C热激35 min后正常培养。)Fig. 9 Thermotolerances assay of yeast harboring pYES2 or pYES2-ZmHsf25 after heat shock at 50° C(A: culture under normal conditions; B: culture under normal conditions after HS at 50° C for 45 min.) |
图10
Fig. 10
| Figure OptionViewDownloadNew Window | |
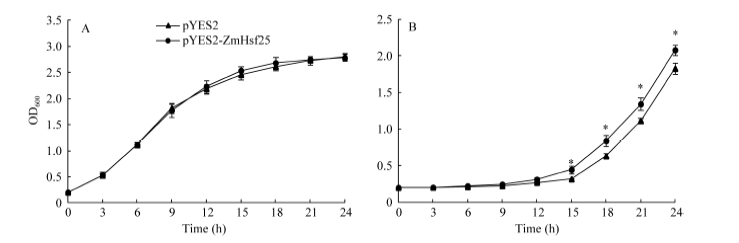 | 图10 50° C热激处理后转基因酵母生长速率比较(A: 正常培养; B: 50° C热激15 min后正常培养。)Fig. 10 Comparison of growth ratios between yeast harboring pYES2 and pYES2-ZmHsf25 after heat shock at 50° C(A: culture under normal conditions; B: culture under normal conditions after HS at 50° C for 45 min.) |
3 讨论植物热激转录因子属于多基因家族, 2011年, Lin等[20]通过生物信息学推测, 玉米有25个家族成员, 2016年, 数目增加为30个[3], 其中B族有7个。关于玉米个体Hsf功能研究报道不多。本实验室在前期研究玉米A1亚族ZmHsf06的基础上[19, 24], 从玉米幼叶中克隆到多个Hsf, 通过结构分析并与前人推测结果比对, 其中一个属于B族, 为ZmHsf25。进一步分析表明, 该基因完整编码序列为957个核苷酸, 编码318个氨基酸残基。蛋白质结构含有完整的DNA结合结构域, 同时含有核定位信号和核输出信号序列, 但不含激活结构域。同源分析表明, ZmHsf25蛋白与高粱Sb060g025710、谷子SiHSFB- 2a-like和水稻OSIGBa0103M18.7的相似性分别为92%、81%和77%。拟南芥、番茄和大豆HsfB1的C末段均含有一段保守序列, 即11个氨基酸GEGLKLFGVWL的BRD抑制域, 为基因HsfB1抑制热诱导基因表达所必需[25, 26]。但是, 作为HsfA1共激活因子, 番茄HsfB1依赖于C末段一段类组氨酸序列(GRGKMMK), 该序列为植物CBP/HAC1的结合位点[27]。可见, C末段氨基酸序列不同导致基因功能的差异性。比对表明, ZmHsf25与番茄和拟南芥HsfB1相似性较低, C末段既不含11个氨基酸的保守序列, 也不含类组氨酸序列。那么, ZmHsf25的功能如何有待深入研究。
因B族Hsf不含激活结构域, 早期研究认为其主要起抑制作用[9, 10]。随着研究的深入, 越来越多的研究发现, B族Hsf受多种逆境胁迫诱导表达, 从正反方面, 参与植株耐热、抗旱和抗病等多种生物和非生物胁迫及生长发育过程[10, 11, 12, 13, 28]。非胁迫条件下, 拟南芥HsfB1和HsfB2b定位在细胞核, 直接抑制HsfA2、HsfA7a、HsfB1和HsfB2b的表达[10]。从本研究基因定位和表达结果来看, 正常生长条件下, ZmHsf25定位在细胞核, ZmHsf25表达量在多数组织中不高, 但在幼嫩的花粉中显著高于在其他组织中。逆境胁迫下, 基因呈现不同表达谱, 热胁迫能显著上调根系和叶片ZmHsf25的表达, 水杨酸和H2O2处理对根系和叶片ZmHsf25的表达均表现为下调。如果水杨酸和H2O2预处理后再进行热激, 二者均能显著上调ZmHsf25的表达, 表明水杨酸和H2O2在热激条件下才能诱导ZmHsf25大幅度表达。前人研究认为, 水杨酸介导植物耐热性尤其是植株获得耐热性过程[29, 30], 水杨酸可在上游调控拟南芥AtHsfA2的表达, 该过程需要借助H2O2信号途径, 同时依赖相关热激蛋白基因的表达[31]。这间接表明ZmHsf25可能是水杨酸热激信号转导过程中的下游组分之一, 初步显示出ZmHsf25在植株花粉发育和耐热过程中的调控作用。正常条件下, 水杨酸下调根系ZmHsf25表达, 可能是水杨酸处理浓度较大所致。作为信号分子, 水杨酸在植株体内的存在是极其微量的。
自20世纪80年代后期酵母Hsf被首次克隆以来, 越来越多动植物物种的Hsf基因被相继获得和注释。酵母中只有1个Hsf, 相对简单, 以酵母为转化系统来验证Hsf的功能越来越得到应用[32]。本研究通过转化酵母发现, 正常生长条件下, 转基因酵母和正常对照长势没有明显差异; 经50℃热激处理45 min后, 转基因酵母细胞和转化空载体对照的生长速度均受到抑制, 但是前者的受抑制程度明显小于后者, 表明ZmHsf25能在酵母细胞中被诱导表达, 且能显著提高酵母细胞的耐热性, 进一步验证了ZmHsf25的耐热性调控功能。目前正在通过转基因拟南芥植株对ZmHsf25的耐热性功能深入研究。
4 结论玉米B族热激转录因子基因ZmHsf25的完整编码序列全长957 bp, 编码318个氨基酸残基。蛋白质序列含有DNA结合结构域、核定位信号序列和核输出信号序列。ZmHsf25与高粱Sb06g025710的相似性最高, 达92%。正常生长条件下, ZmHsf25在玉米多个组织器官中表达, 幼嫩花粉中表达量最高。42℃热胁迫能显著上调根系和叶片中ZmHsf25的表达; 正常条件下, 水杨酸和H2O2处理下调根系和叶片ZmHsf25表达, 而热激条件下却表现为显著上调。ZmHsf25定位在细胞核。转ZmHsf25酵母具有较强的耐热性。ZmHsf25可能主要参与玉米植株花粉发育和热胁迫响应过程, 推测为水杨酸热激信号转导的下游组分之一。
The authors have declared that no competing interests exist.
作者已声明无竞争性利益关系。The authors have declared that no competing interests exist.
参考文献View Option
原文顺序
文献年度倒序
文中引用次数倒序
被引期刊影响因子
| [1] | |
| [2] | |
| [3] | |
| [4] | |
| [5] | |
| [6] | |
| [7] | |
| [8] | |
| [9] | |
| [10] | |
| [11] | |
| [12] | |
| [13] | |
| [14] | |
| [15] | |
| [16] | |
| [17] | |
| [18] | |
| [19] | |
| [20] | |
| [21] | |
| [22] | |
| [23] | |
| [24] | |
| [25] | |
| [26] | |
| [27] | |
| [28] | |
| [29] | |
| [30] | |
| [31] | |
| [32] |

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}