关键词:普通小麦近缘种; 低分子量麦谷蛋白亚基; Glu-A3基因; 等位变异 Isolation and Characterization of LMW-GS Glu-A3in Common Wheat Related Species DONG Xue1, LIU Meng1, ZHAO Xian-Lin2,*, FENG Yu-Mei1, YANG Yan1,* 1 The functional Laboratory of Plant Biotechnology, College of Life Sciences, Inner Mongolia Agricultural University, Huhhot 010018, China
2Wheat Research Institute, Henan Academy of Agricultural Sciences / Henan Key Laboratory of Wheat Biology / National Engineering Laboratory for Wheat / Key Laboratory of Wheat Biology and Genetic Breeding in Central Huang-Huai Region, Ministry of Agriculture, Zhengzhou 450002, China
Fund:This study was supported by the National Natural Science Foundation of China (31271726), the Natural Science Foundation of Inner Mongolia (2014MS0338), the Excellent Young Scientist Foundation of Inner Mongolia Agricultural University (2014XYQ-18), and the Chunhui Project funded by the Ministry of Education (Z2009-1-01060). AbstractLow-molecular-weight glutenin subunits (LMW-GS) account for about one-third of wheat seed storage proteins and have great effects on dough extensibility and food processing quality. Common wheat relative species are important genetic resources for wheat improvement. In this study, the specific primers of Glu-A3 genes were used to identify the allelic variations of GluA3-1, GluA-2, and GluA3-3 from nine accessions of Triticum dicoccoides, T. dicoccum, T. durum, and T. boeoricum. A total of 15 new haplotypes were identified, including five in GluA3-1, five in GluA3-2, and five in GluA3-3. Among them, two haplotypes each contained nine cysteines that might be good for improvement of wheat quality. The results about Glu-A3 genes in wheat further made clear composition of LMW-GS and provide a basis for selection of parents in wheat quality breeding.
Keyword:Common wheat related species; LMW-GS; Glu-A3gene; Allelic variation Show Figures Show Figures
表1Glu-A3位点基因特异性引物 Table 1 Locus-specific primers used for amplifyingGlu-A3 genes
目标基因 Target gene
引物 Primer
引物序列 Primer sequence (5′ → 3′ )
参考基因 Reference gene
引物位置 Primer location
目的片段大小 Fragment size (bp)
退火温度 Anneal temp. (° C)
GluA3-1
LA1F
AAACAGAATTATTAAAGCCGG
EU189087
662
~1500
57
LA1R
CAGGCAAGACAAACTTATGAA
EU189087
2198
GluA3-2
LA2F
CATCTCTGATCCATCCAAACAT
AB062868
50
~1100
57
LA2R
GTGATCCAACTATATATTACTAGAC
FJ441117
1007
GluA3-3
LA3F
TTCAGATGCAGCCAAACAA
EF426565
71283
~1400
58
LA3R
CAAACTTATGAACTTGCAAGG
EF426565
72669
Primer location counted from the first nucleotide of the available gene fragments. Primer sequences are from Wang [23]. 引物位置从参考基因的第一个核苷酸计算。引物序列来自王林海[23]。
表1Glu-A3位点基因特异性引物 Table 1 Locus-specific primers used for amplifyingGlu-A3 genes
附图1Glu-A3位点推导氨基酸序列比对Supplementary figure 1 Alignments of deduced amino-acid sequences on Glu-A3 loci (A): AP1-7与5个GluA3-1单元型推导氨基酸的序列比对; (B): AP2-3与5个GluA3-2单元型推导氨基酸的序列比对; (C): AP3-6与 5个GluA3-3单元型推导氨基酸的序列比对。带下画线的加粗体字母表示半胱氨酸残基, 灰色背景显示突变位点。黑色和白色倒三角分别指示N末端保守区和重复区的起始点; 箭头指表示C末端的3个亚区, 包括半胱氨酸富集区、谷酰胺富集区和末端保守区的起始点。 (A): Alignments of deduced amino-acid sequences (AP1-7) of oneGluA3-1 haplotype and those (AP1-8 to AP1-12) of five GluA3-1 genes; (B): Alignments of deduced amino-acid sequences (AP2-3) of oneGluA3-2 haplotype and those (AP2-7 to AP2-11) of five GluA3-2 genes; (C): Alignments of deduced amino-acid sequences (AP3-6) of one GluA3-3 haplotype and those (AP3-9 to AP3-13) of five GluA3-3 genes. The positions of the cysteine residues are bold and underlined, and mutation loci are shadowed. Filled nabla the beginning of short N-terminal conserved region; open nabla shows the beginning of N-terminal repetitive domain; down arrow shows the beginning of the three subregions of C-terminal part of the protein, including a cysteine-rich region, a glutamine-rich region and shows the final conserved part of proteins, respectively.
图1 3对Glu-A3位点基因特异性引物在普通小麦近缘种中的PCR扩增产物电泳检测Fig. 1 PCR products of Glu-A3 amplified with three gene-specific primers in common wheat relative species A: LA1F/LA1R扩增产物(1438~1537 bp); B: LA2F/LA2R扩增产物(1070~1100 bp); C: LA3F/LA3R扩增产物(1348~1420 bp)。M: DL2000 marker; 1: 野生二粒小麦DS1; 2: 野生二粒小麦DS3; 3: 野生二粒小麦15; 4: 栽培二粒小麦DM2; 5: 栽培二粒小麦DM3; 6: 硬粒小麦Langdon; 7: 野生一粒小麦B01; 8: 野生一粒小麦B03; 9: 野生一粒小麦B04。 A: PCR products amplified with the primer set LA1F/LA1R (1438-1537 bp); B: PCR products amplified with the primer set LA2F/LA2R (1070-1100 bp); C: PCR products amplified with the primer set LA3F/LA3R (1348-1420 bp). M: DL2000 marker; 1: T. dicoccoides DS1; 2: T. dicoccoidesDS3; 3: T. dicoccoides 15; 4: T. dicoccum DM2; 5: T. dicoccumDM3; 6: T. durum Langdon; 7: T. boeoricum accession B01; 8: T. boeoricum B03; 9: T. boeoricumB04.
表2 本研究鉴定的Glu-A3位点新单元型与GenBank中相关序列的同源性比较 Table 2 Similarity comparison between novel Glu-A3 haplotypes identified in this study and related sequences deposited in GenBank (%)
Gene
GluA3-18
GluA3-19
GluA3-110
GluA3-111
GluA3- 112
GluA3-27
GluA3- 28
GluA3- 29
GluA3- 210
GluA3- 211
GluA3-39
GluA3-310
GluA3-311
GluA3-312
GluA3-313
GluA3-11
98.5
99.5
98.6
99.3
97.2
87.6
87.0
87.6
87.5
87.7
95.6
95.3
95.0
94.6
94.8
GluA3-12
98.7
99.6
98.8
99.5
97.3
87.1
86.5
87.1
87.0
87.2
95.2
95.1
94.8
94.4
94.5
GluA3-13
98.5
99.5
98.7
99.4
97.2
87.7
87.1
87.7
87.6
87.8
95.6
95.3
95.0
94.6
94.8
GluA3-14
96.8
97.2
96.7
97.0
99.0
86.8
86.3
87.0
86.9
87.1
94.6
94.1
94.1
93.5
93.8
GluA3-15
99.2
98.1
97.9
97.8
96.5
86.6
85.9
86.5
86.4
86.6
94.5
94.1
93.8
93.4
93.7
GluA3-16
97.7
98.2
97.7
97.9
96.6
86.2
85.8
86.5
86.4
86.5
94.6
94.5
94.3
93.9
93.9
GluA3-17
99.9
98.6
98.3
98.3
96.7
86.7
86.0
86.6
86.5
86.7
94.9
94.3
94.2
93.8
94.0
GluA3-21
84.6
85.4
85.0
85.1
84.7
99.3
99.2
99.6
99.5
99.7
86.2
85.5
85.7
85.2
85.5
GluA3-22
84.6
85.4
85.0
85.2
84.8
99.3
99.2
99.6
99.5
99.7
86.2
85.5
85.7
85.2
85.5
GluA3-23
84.3
85.1
84.7
84.8
84.4
99.1
99.1
99.4
99.3
99.5
85.9
85.1
85.3
84.8
85.2
GluA3-24
84.7
85.4
85.0
85.1
84.9
99.1
98.9
99.4
99.3
99.5
86.0
85.3
85.5
84.9
85.3
GluA3-31
93.9
94.6
94.2
94.4
94.4
85.8
85.7
86.1
86.0
86.4
97.1
99.2
97.0
96.3
96.8
GluA3-32
93.6
94.2
93.8
94.0
94.0
85.3
85.2
85.6
85.5
85.9
96.8
98.9
96.7
96.0
96.5
GluA3-33
93.5
93.8
93.8
93.5
93.6
85.5
85.2
85.7
85.6
85.9
99.4
96.5
97.9
98.2
98.3
GluA3-34
93.7
94.2
94.1
94.0
93.7
86.0
85.8
86.3
86.2
86.5
97.0
98.4
97.0
96.1
96.8
GluA3-35
93.5
94.1
93.8
93.9
93.9
85.3
85.2
85.6
85.5
85.9
96.8
98.9
96.7
96.0
96.5
GluA3-36
93.5
94.2
93.9
94.0
93.7
86.0
85.8
86.3
86.2
86.5
96.9
98.3
96.7
95.8
96.6
DQ630443
95.1
95.6
94.9
95.5
96.9
80.3
79.6
80.2
80.1
80.4
93.0
92.5
92.6
92.3
91.4
AB062877
94.7
95.2
94.6
95.2
97.1
80.0
79.4
79.9
79.8
80.0
92.8
92.9
92.6
92.1
91.4
AB062878
95.2
95.7
95.1
95.6
97.2
80.0
79.4
79.9
79.8
80.0
93.2
93.1
92.8
92.4
91.7
AY453157
96.5
97.2
96.4
97.0
99.1
70.3
69.5
69.6
70.2
70.1
92.7
93.0
91.9
92.1
91.9
AY453159
97.6
98.7
97.5
98.4
96.7
67.7
67.8
68.5
68.4
68.5
91.3
89.4
91.2
90.2
90.4
X07747
97.5
98.0
97.3
97.8
96.3
79.0
78.2
78.8
78.7
78.9
90.5
89.0
91.3
89.3
90.3
AJ293097
99.2
97.9
97.4
97.8
96.2
72.6
72.2
72.9
72.8
72.9
90.6
88.7
90.3
89.7
89.7
AY146587
99.7
98.3
97.8
98.2
96.5
80.2
79.8
80.4
80.3
80.5
90.2
88.7
91.0
88.9
89.7
AY453154
98.3
99.3
98.2
99.2
97.3
71.4
71.2
71.8
71.7
71.7
91.0
89.8
90.6
90.6
90.3
AY453155
98.6
99.6
98.4
99.5
97.5
73.0
72.7
73.3
73.2
73.3
92.0
90.5
91.5
91.1
91.0
AY453156
98.4
99.4
98.3
99.3
97.4
71.5
71.3
71.9
71.8
71.8
91.1
89.9
90.7
90.7
90.4
AY453158
99.0
97.9
97.4
97.7
96.5
70.5
70.3
70.8
70.7
70.8
91.4
89.9
91.1
90.4
90.4
AY453160
99.8
98.5
97.8
98.4
96.7
73.4
73.1
73.7
73.6
73.7
91.2
89.7
91.1
90.2
90.3
AY542896
99.8
98.5
97.9
98.4
96.8
80.2
79.8
80.4
80.3
80.5
93.9
92.4
93.7
92.6
93.3
DQ517534
98.1
96.6
96.0
96.5
94.7
75.4
75.0
75.6
75.5
75.6
89.3
87.3
89.1
88.5
88.6
DQ519083
98.6
97.3
96.7
97.2
95.4
76.3
75.9
76.5
76.4
76.5
90.1
88.9
90.1
89.5
89.6
DQ519084
96.3
94.8
94.1
94.7
92.7
73.6
73.2
73.8
73.7
73.8
87.2
85.2
87.0
86.4
86.5
DQ519085
98.3
97.2
96.7
97.0
95.7
75.1
74.6
75.2
75.2
75.1
90.6
88.7
90.4
89.7
89.9
EU189087
99.9
98.5
98.0
98.4
96.7
80.2
79.8
80.4
80.3
80.5
90.5
89.0
91.1
89.2
90.0
AB062868
75.3
76.0
75.6
76.1
75.9
99.0
98.8
99.2
99.2
99.4
81.0
80.4
80.6
79.9
80.4
X84959
71.5
72.1
71.5
72.1
73.6
95.4
95.6
95.6
95.6
96.1
72.5
70.1
71.5
71.7
71.3
AJ293098
92.7
94.4
93.8
94.0
93.0
73.4
73.7
74.2
74.1
74.4
95.1
97.3
94.7
94.4
92.9
DQ217663
98.9
97.8
97.1
97.7
96.2
78.0
77.5
78.1
78.0
78.1
90.0
88.6
90.6
88.7
89.3
EF426565
92.2
92.9
92.5
92.7
92.2
76.9
77.0
77.5
77.4
77.6
96.7
98.1
96.5
95.7
96.5
FJ441117
77.1
77.4
76.7
77.3
77.5
97.5
97.6
97.7
97.7
98.0
78.5
77.6
77.8
77.2
77.1
不考虑序列长度。加粗体表示假基因。Irrespective of the sequence length. Pseudogenes are in bold font.
表2 本研究鉴定的Glu-A3位点新单元型与GenBank中相关序列的同源性比较 Table 2 Similarity comparison between novel Glu-A3 haplotypes identified in this study and related sequences deposited in GenBank (%)
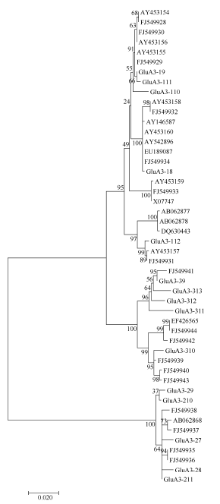
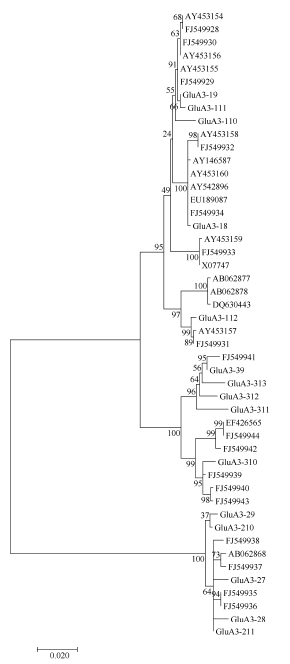
图2 来源于普通小麦及其近缘种中48个Glu-A3基因的进化树分析(MEGA 6.0, NJ法)Fig. 2 Phylogenetic tree of Glu-A3 genes in common wheat and related species (software MEGA version 6.0 with Neighbour Joining)
表3 3个Glu-A3基因的15个新单元型(下对角线)及其推导氨基酸(上对角线)序列的相似性比较 Table 3 Similarity comparison of 15 new haplotypes from threeGlu-A3 genes (below diagonal) and their deduced amino-acid sequences (above diagonal) (%)
Gene
GluA3-18
GluA3-19
GluA3-110
GluA3-111
GluA3-112
GluA3-27
GluA3-28
GluA3-29
GluA3-210
GluA3-211
GluA3-39
GluA3-310
GluA3-311
GluA3-312
GluA3-313
GluA3-18
100.0
97.3
―
96.6
95.8
―
76.3
76.5
76.8
77.1
―
―
―
90.1
―
GluA3-19
98.5
100.0
―
99.2
96.8
―
79.8
79.9
80.3
80.6
―
―
―
91.0
―
GluA3-110
97.9
98.5
100.0
―
―
―
―
―
―
―
―
―
―
―
―
GluA3-111
98.3
99.5
98.1
100.0
96.0
―
79.4
79.6
79.9
80.2
―
―
―
90.7
―
GluA3-112
96.7
97.2
96.4
97.0
100.0
―
78.4
78.9
78.9
79.3
―
―
―
90.2
―
GluA3-27
87.5
87.9
87.1
87.8
87.9
100.0
―
―
―
―
―
―
―
―
―
GluA3-28
86.8
87.3
86.5
87.2
87.5
98.9
100.0
98.7
98.7
99.0
―
―
―
78.5
―
GluA3-29
87.3
87.8
87.0
87.7
88.0
99.7
99.2
100.0
100.0
99.7
―
―
―
78.9
―
GluA3-210
87.2
87.7
86.9
87.6
87.9
99.8
99.1
99.8
100.0
99.7
―
―
―
79.3
―
GluA3-211
87.5
88.0
87.2
87.9
88.2
99.5
99.4
99.7
99.7
100.0
―
―
―
79.3
―
GluA3-39
94.1
94.3
94.0
94.0
94.3
85.0
84.8
85.5
85.4
85.1
100.0
―
―
―
―
GluA3-310
93.3
93.8
93.3
93.3
93.2
84.5
84.4
84.9
84.8
84.8
97.2
100.0
―
―
―
GluA3-311
93.2
93.5
93.3
93.1
93.2
84.8
84.5
85.2
85.1
84.8
98.2
96.9
100.0
―
―
GluA3-312
93.0
93.4
92.9
93.0
93.0
83.8
83.5
84.2
84.1
84.0
98.5
96.3
97.7
100.0
―
GluA3-313
93.3
93.6
93.3
93.2
93.4
83.7
83.5
84.1
84.0
83.8
98.8
96.8
97.7
98.1
100.0
New haplotypes are below diagonal; new haplotypes deduced amino-acid sequences are above diagonal; irrespective of the sequence length. Pseudogenes are in bold font. 下对角线为新单元型, 上对角线为推导氨基酸, 不考虑序列长度。加粗体表示假基因。
表3 3个Glu-A3基因的15个新单元型(下对角线)及其推导氨基酸(上对角线)序列的相似性比较 Table 3 Similarity comparison of 15 new haplotypes from threeGlu-A3 genes (below diagonal) and their deduced amino-acid sequences (above diagonal) (%)
LöveÁ. Conspectus of the Triticeae. Feddes Rep, 1984, 95: 425-521[本文引用:1]
[2]
LuoC, Griffin WB, BranlardG, Mcneil DL. Comparison of low- and high molecular-weight wheat glutenin allele effects on flour quality. Theor Appl Genet, 2001, 102: 1088-1098[本文引用:3]
[3]
Zhen SM, Han CX, Ma CY, Gu AQ, ZhangM, Shen XX, Li XH, Yan YM. Deletion of the low-molecular-weight glutenin subunit allele Glu-A3a of wheat( Triticum aestivumL . ) significantly reduces dough strength and breadmaking quality. BMC Plant Biol, 2014, 14: 367[本文引用:1]
[4]
McIntosh RA, YamazakiY, Devos KM, DubcovskyJ, Rogers WJ, AppelsR. Catalogue of gene symbols for wheat. Proceedings of the 10th international wheat genetics symposium, Paestum, Italy, 2003[本文引用:2]
[5]
Liu CY, Shepherd KW. Inheritance of B subunits of glutenin and ω-and γ-gliadins in tetraploid wheats. Theor Appl Genet, 1995, 90: 1149-1157[本文引用:1]
[6]
SreeramuluG, Singh NK. Genetic and biochemical characterization of novel low molecular weight glutenin subunits in wheat. Genome, 1997, 40: 41-48[本文引用:1]
[7]
D'OvidioR, Masci S. The low-molecular-weight glutenin subunits of wheat gluten. J Cereal Sci, 2004, 39: 321-339[本文引用:2]
[8]
Luo GB, Zhang XF, Zhang YL, Yang WL, Li YW, Sun JZ, Zhan KH, Zhang AM, Liu DC. Composition, variation, expression and evolution of low-molecular-weight glutenin subunit genes in Triticum urartu. BMC Plant Biol, 2015, 15: 68[本文引用:2]
Wang YP, Zhen SM, Luo NN, Han CX, Lu XB, Li XH, Xia XC, He ZH, Yan YM. Low molecular weight glutenin subunit gene Glu-B3h confers superior dough strength and breadmaking quality in wheat( Triticum aestivumL . ). Sci Rep, 2016, 6: 27182[本文引用:1]
[11]
Si HQ, Zhao ML, ZhangX, Yao GuoL, Sun GL, Ma CX. Cloning and characterization of low-molecular-weight glutenin subunit alleles from Chinese wheat land races ( Triticum aestivumL. ). Sci World J, 2014, 2014: 371045[本文引用:2]
[12]
Wang LH, Zhao XL, He ZH, MaW, AppelsR, Peña RJ, Xia XC. Characterization of low-molecular-weight glutenin subunit Glu-B3 genes and development of STS markers in common wheat( Triticum aestivum L. ). Theor Appl Genet, 2009, 118: 525-539[本文引用:4]
[13]
Zhao XL, YangY, He ZH, Lei ZS, MaW, Sun QX, Xia XC. Characterization of novel LMW-GS genes at Glu-D3 locus of chromosome 1D in Aegilops tauschii. Hereditas, 2008, 145: 238-250[本文引用:3]
[14]
程西永, 吴少辉, 李海霞, 董中东, 任妍, 詹克慧, 许海霞. 小麦高、低分子量麦谷蛋白亚基对品质性状的影响. 麦类作物学报, 2014, 34: 482-488Cheng XY, Wu SH, Li HX, Dong ZD, RenY, Zhan KH, Xu HX. Effects of HMW and LMW glutenin subunits on wheat quality traits. J Triticeae Crops, 2014, 34: 482-488 (in Chinese with English abstract)[本文引用:1]
[15]
ItoM, FushieS, Funatsuki WM, Ikeda TM, NishioZ, NagasawaK, TabikiT, YamauchiH. Effect of allelic variation in three glutenin loci on dough properties and bread-making qualities of winter wheat. Breed Sci, 2011, 61: 281-287[本文引用:1]
[16]
JinH, ZhangY, LiG, MuP, FanZ, XiaX, HeZ. Effects of allelic variation of HMW-GS and LMW-GS on mixograph properties and Chinese noodle and steamed bread qualities in a set of Aroona near-isogenic wheat lines. J Cereal Sci, 2013, 57: 146-152[本文引用:1]
[17]
ZhangX, JinH, ZhangY, LiuD, LiG, XiaX, ZhangA. Composition and functional analysis of low-molecular-weight glutenin alleles with Aroona near-isogenic lines of bread wheat. BMC Plant Biol, 2011, 12: 1-16[本文引用:1]
[18]
Qin LM, LiangY, Yang DZ, SunL, Xia GG, Liu SW. Novel LMW glutenin subunit genes from wild emmer wheat ( Triticum turgidumssp. dicoccoides) in relation to Glu-3 evolution. Dev Genes Evol, 2015, 225: 31-37[本文引用:1]
ZhangW, Gianibelli MC, Rampling LR, Gale KR. Characterisation and marker development for low molecular weight glutenin genes from Glu-A3 alleles of bread wheat( Triticum aestivum L. ). Theor Appl Genet, 2004, 108: 1409-1419[本文引用:1]
赵曼丽. 低分子量麦谷蛋白亚基基因的克隆及序列分析. 安徽农业大学硕士学位论文, 安徽合肥, 2014Zhao ML. Cloning and Sequence Analysis of Low Molecular Weight Glutein Subunit Gene. MS Thesis of Anhui Agricultural University, Hefei, China, 2014 (in Chinese with English abstract)[本文引用:1]
[23]
王林海. 普通小麦及其近缘种低分子量麦谷蛋白基因克隆与STS标记开发. 中国农业科学院博士学位论文, 北京, 2009Wang LH. Characterisation of Low-Molecular-Weight Glutenin Genes and Development of STS Markers in Common Wheat and Its Relatives. PhD Dissertation of Chinese Academy of Agricultural Sciences, Beijing, China, 2009 (in Chinese with English abstract)[本文引用:7]
[24]
Gale KR, MaW, ZhangW, RamplingL, Hill AS, AppelsR, MorrisP, MorrelM. Simple high-throughput DNA markers for genotyping in wheat. In: Eastwood R, Hollamby G, Rathjen T, Gororo N, eds. Proceedings of 10th Australian Wheat Breeding Assembly. Mildura VIC: Wheat Breeding Society of Australia, 2001. pp 26-31[本文引用:1]
[25]
StepienL, Holubec V. ChelkowskiJ. Resistance genes in wild accessions of Triticeae-inoculation test and STS marker analyses. Theor Appl Genet, 2002, 43: 423-435[本文引用:1]
[26]
Qin LM, LiangY, Yang DZ, SunL, Xia GM, Liu SW. Novel LMW glutenin subunit genes from wild emmer wheat ( Triticum turgidumssp. dicoccoides) in relation to Glu-3evolution. Dev Genes Evol, 2015, 225: 1-7[本文引用:1]
[27]
LongH, Wei YM, Yan ZH, BaumB, NevoE, Zhen YL. Classification of wheat low-molecular-weight glutenin subunit genes and its chromosome assignment by developing LMW-GS group-specific primers. Theor Appl Genet, 2005, 111: 1251-1259[本文引用:1]
[28]
Zhao HX, Wang RJ, Guo AG, Hu SW, Sun GL. Development of primers specific for LMW-GS genes located on chromosome 1D and molecular characterization of a gene from Glu-D3complex locus in breed wheat. Hereditas, 2004, 141: 193-198[本文引用:2]
[29]
XuH, Wang RJ, ShenX, Zhao YL, Sun GL, Zhao HX, Guo AG. Functional properties of a new low-molecular-weight glutenin subunit gene from a bread wheat cultivar. Theor Appl Genet, 2006, 113: 1295-1303[本文引用:1]
 , 刘梦
, 刘梦




{kind=link}
{kind=link}
{kind=link}