关键词:稻瘟病; 种质资源; 自然诱发; 抗谱; 优异抗源 Identification and Evaluation of Blast Resistance for Resequenced Rice Core Collections LI Xu-Sheng1,**, XIANG Xiao-Jiao2,**, SHEN Cong-Cong2, YANG Long-Wei1,*, CHEN Kai3, WANG Xiao-Wen1, QIU Xian-Jin1, ZHU Xiao-Yuan4, XING Dan-Ying1, XU Jian-Long2,3,* 1 Hubei Collaborative Innovation Centre for Grain Industry, Yangtze University, Jingzhou 434025, China
2 Institute of Crop Science, Chinese Academy of Agricultural Sciences, Beijing 100081, China
3Agricultural Genomics Institute at Shenzhen, Chinese Academy of Agricultural Sciences, Shenzhen 518210, China
4 Institute of Rice Research, Guangdong Academy of Agricultural Sciences, Guangzhou 510640, China
Fund:This study was supported by National High-Tech Research & Development Plan (863 program) (2014AA10A601), Open Fund of Hubei Collaborative Innovation Centre for Grain Industry (2015MS010, LXT-16-06, LXT-17-02), the Shenzhen Peacock Plan (20130415095710361), Collaboration Project of Hubei Academy of Agricultural Sciences (2015H200014), and Open Fund of Research Centre of Ministry of Education for Wetland Ecology & Agronomy Application (KF201403). AbstractRice blast is one of the key factors that restrict rice yield, and screening blast resistance resources is a basis work for mining blast resistance genes and breeding resistant varieties. A set of 1217 accessions selected from 3000 (3K) re-sequenced rice core collection were comprehensively evaluated for blast resistance in two natural disease nurseries at Lianghe and Bajiao in Enshi area and agronomic traits in disease-free field as well as resistance spectrum of resistant accessions at seedling stage. There were significant differences in response of different accessions to rice blast. Among them, a total of 144 accessions with blast resistance at different growing stages were obtained. Thirty-four accessions with relatively high comprehensive blast resistance were inoculated with 30 diverse isolates, showing that 17 accessions with a broad resistance spectrum (≥70%). According to the investigation of agronomic traits, most resistant accessions had high plant height, low grain yield per plant and poor agronomic traits. Finally, seven accessions with high blast resistance, broad resistant spectrum and relatively good agronomic traits were selected, including IRGA411-1-6-1F-A, YJ 30, Jinzao 47, Quanzhen 10, YN 1353-3, Yunjing 23, and IRAT1047, were identified. Those resistant accessions can be used in mining resistance gene and variety improvement for blast resistance.
Keyword:Rice blast; Germplasm; Natural infestation; Resistance spectrum; Excellent resistance resource Show Figures Show Figures
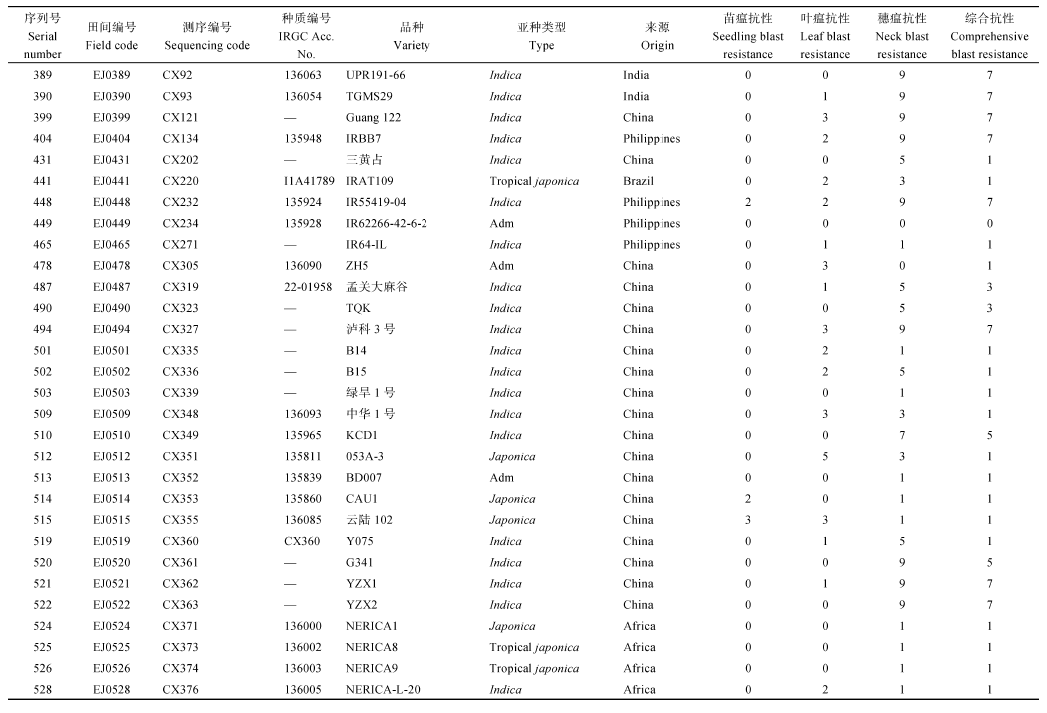
表4 病圃综合抗性强和田间表现较好材料的稻瘟病病情 Table 4 Blast incidence of materials with good blast resistance and field performance in the nursery
田间号 Field code
品种名称 Variety name
平均苗瘟 病级 Average seedling blast grade
平均叶瘟 病级 Average leaf blast grade
平均 穗瘟率 Average neck blast rate (%)
最高苗瘟 病级 Maximum seedling blast grade
最高叶瘟 病级 Maximum leaf blast grade
最高穗 瘟率 Maximum neck blast rate (%)
最高综合 指数 Maximum comprehensive index
最高综合 病级 Maximum comprehensive grade
EJ0235
IRGA411-1-6-1F-A
0.0
0.0
3.5
0
0
6
1.25
1
EJ0270
IRAT266
0.5
2.8
1.50
1
4
4
1.75
1
EJ0441
IRAT109
0.0
0.8
5.50
0
2
10
1.75
1
EJ0465
IR64-IL
0.0
0.5
0.25
0
1
1
0.50
1
EJ0512
053A-3
0.0
2.5
6.8
0
5
10
2.00
1
EJ0513
BD007
0.0
0.0
0.50
0
0
1
0.25
1
EJ0514
CAU1
1.0
0.0
1.8
2
0
4
0.75
1
EJ0515
云陆102 Yunlu 102
2.5
1.5
0.75
3
3
2
1.50
1
EJ0537
YJ30
4.0
4.5
1.25
4
5
2
2.00
1
EJ0546
611
0.0
0.0
2.00
0
0
4
0.75
1
EJ0611
金早47 Jinzao 47
0.0
0.0
0.00
0
0
0
0.00
0
EJ0616
泉珍10号Quanzhen 10
0.0
0.0
8.8
0
0
10
1.25
1
EJ0619
密阳46 Milyang 46
4.0
2.0
0.75
4
4
2
1.75
1
EJ0641
SET
4.0
3.0
0.00
4
4
0
1.00
1
EJ0666
YN1353-3
0.0
1.5
0.00
0
3
0
0.75
1
EJ0744
旱277 Han 277
4.5
4.5
5.0
5
5
5
2.00
1
EJ0748
陆引46 Luyin 46
0.0
1.5
1.0
0
3
2
1.50
1
EJ0752
云粳23 Yunjing 23
5.0
4.5
0.00
5
5
0
1.25
1
EJ0790
LOCAL BHAT
4.5
4.8
6.5
5
5
10
2.50
3
EJ0868
B6144-MR-6-0-0
0.0
0.5
0.00
0
1
0
0.25
1
EJ0876
GOOLARAH
5.0
4.3
4.50
5
5
6
2.50
3
EJ0885
IRAT104
0.0
0.0
0.00
0
0
0
0.00
0
EJ0952
越富 Yuefu
0.0
2.0
0.50
0
4
2
1.75
1
EJ0964
巴西糯稻 Baxinuodao
0.0
0.5
3.50
0
1
4
1.00
1
EJ0967
IRAT 104
0.0
1.0
0.00
0
2
0
0.50
1
EJ0973
云陆103 Yunlu 103
3.5
2.3
0.00
4
4
0
1.00
1
EJ1082
MARANHAO BRANCO
4.0
2.3
3.50
4
4
8
2.25
3
EJ1083
MIUDO ROXO
4.0
2.0
3.30
4
4
5
1.75
1
EJ1121
CENTURY PATNA
0.0
0.0
0.00
0
0
0
0.00
0
EJ1137
VALTEJO
0.0
0.0
1.00
0
0
4
0.75
1
EJ1142
167
4.0
2.0
0.00
4
4
0
1.00
1
EJ1203
Ginga
4.0
2.0
0.00
4
4
0
1.00
1
EJ1211
IAC47
0.0
1.0
4.80
0
2
7
1.75
1
EJ1214
IAC165
0.0
0.5
0.50
0
1
1
0.50
1
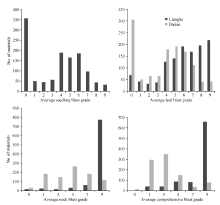
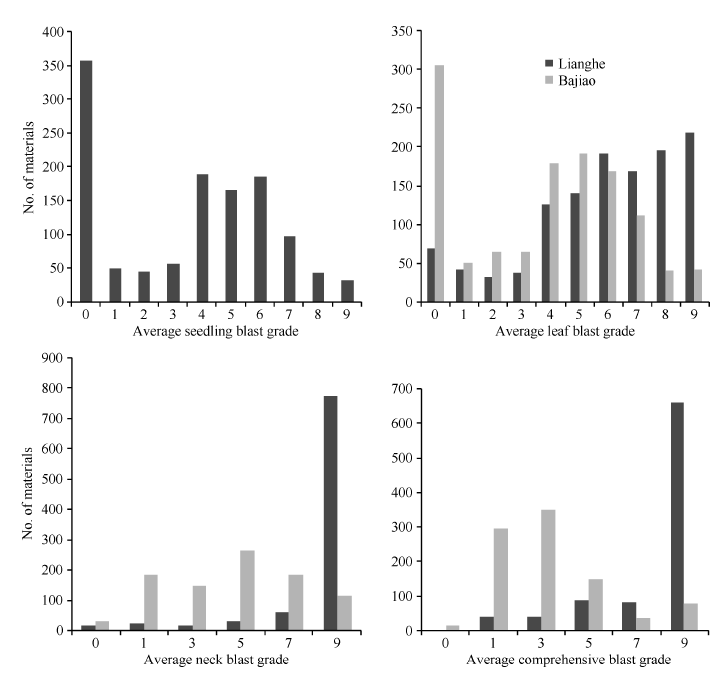
Scores 0-3 stand for resistant while 4-9 for susceptible for seedling and leaf blast; scores 0-1 stand for resistant while 3-9 for susceptible for comprehensive blast grade. 表中苗瘟和叶瘟病0~3级为抗, 4~9级为感; 综合病级0~1级为抗, 3~9级为感。
表4 病圃综合抗性强和田间表现较好材料的稻瘟病病情 Table 4 Blast incidence of materials with good blast resistance and field performance in the nursery
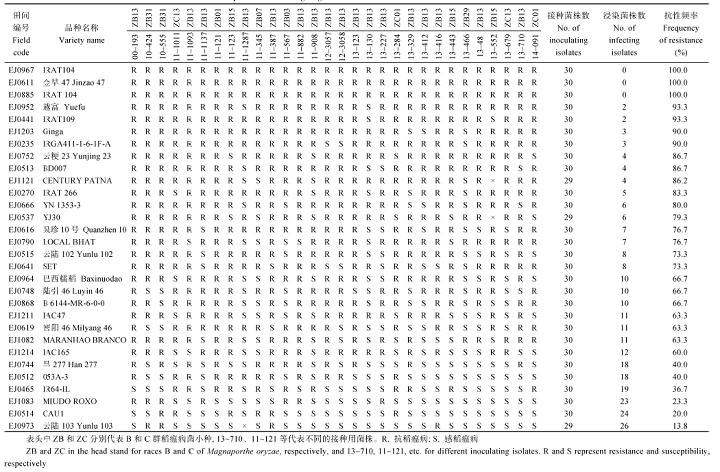
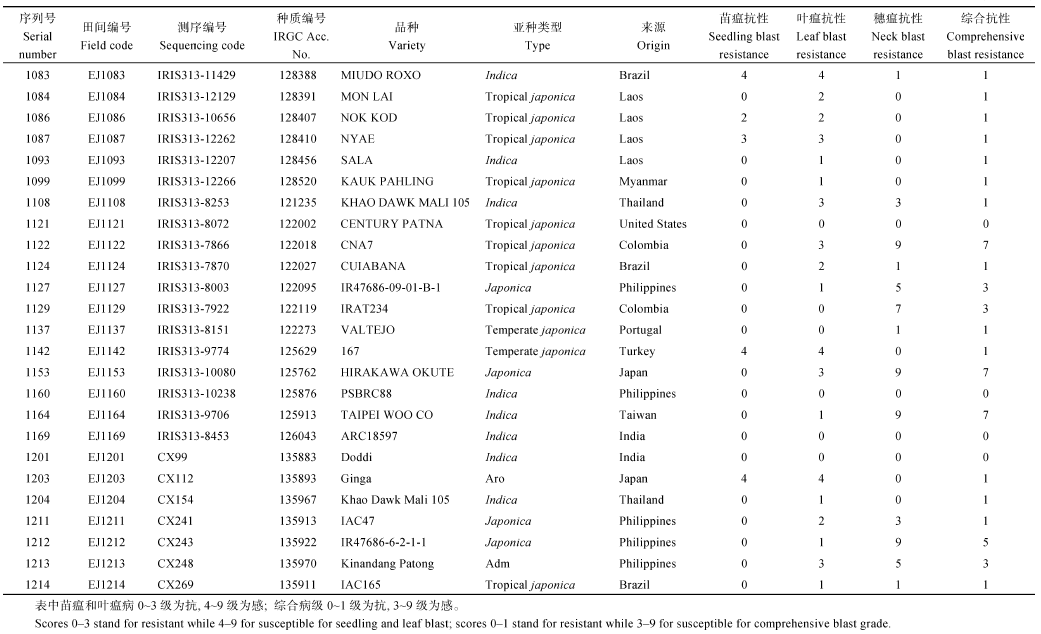
表5 从2个病圃筛选获得的30份抗性材料的苗期抗谱测定 Table 5 Test of resistance spectrum at seedling stage for 30 accessions with llast resistance screened from the two nurseries
表5 从2个病圃筛选获得的30份抗性材料的苗期抗谱测定 Table 5 Test of resistance spectrum at seedling stage for 30 accessions with llast resistance screened from the two nurseries
4 结论对重测序的1217份水稻核心种质资源在恩施两河和芭蕉两个病圃进行稻瘟病抗性自然诱发鉴定, 两病圃的鉴定结果基本一致, 从中获得144份抗苗瘟、叶瘟和穗瘟的抗病种质。选稻瘟病综合抗性较好的34份材料进行30个不同来源的稻瘟病菌株苗期接种鉴定, 筛选出17份抗性频率≥ 70%的广谱抗性材料。鉴定出7份稻瘟病抗性强、抗谱广且农艺性状较好的优异抗源材料IRGA 411-1-6-1F-A、YJ30、金早47、泉珍10号、YN 1353-3、云粳23和IRAT1047, 可作为抗源亲本用于稻瘟病抗性基因挖掘和品种抗稻瘟性改良。 The authors have declared that no competing interests exist.
Hwang BK, Koh YJ, Chung HS. Effects of adult-plant resistance on blast severity and yield of rice. Plant Dis, 1987, 71: 1035-1038[本文引用:1]
[2]
Torres CQ, Teng PS. Path coefficient and regression analysis of the effects of leaf and panicle blast on tropical rice yield. Crop Prot, 1993, 12: 296-302[本文引用:1]
[3]
孙国昌, 杜新法, 陶荣祥, 毛雪琴, 孙漱沅. 水稻稻瘟病防治研究进展和21世纪初研究设想. 植物保护, 2000, 26(1): 33-35Sun GC, Du XF, Tao RX, Mao XQ, Sun SY. Progress in controlling rice blast disease and its prospect in early 21st century. Plant Prot, 2000, 26(1): 33-35 (in Chinese)[本文引用:1]
[4]
Yu ZH, Mackill DJ, Bonman JM, Tanksley SD. Tagging genes for blast resistance in rice via linkage to RFLP markers. Theor Appl Genet, 1991, 81: 471-476[本文引用:1]
[5]
JiaY, Mcadams SA, Bryan GT, Hershey HP, ValentB. Direct interaction of resistance gene and a virulence gene products confers rice blast resistance. EMBO J, 2000, 19: 4004-4014[本文引用:1]
Zeigler RS, Cuoc LX, Scott RP, Bernardo MA, Chen DH, ValentB, Nelson RJ. The relationship between lineage and virulence in Pyricularia grisea in the Philippines. Phytopathology, 1995, 85: 443-451[本文引用:1]
[8]
Turnbull LA, Levine JM, Fergus A J F, Petermann J S. The pi40 gene for durable resistance to rice blast and molecular analysis of pi40-advanced backcross breeding lines. Phytopathology, 2009, 99: 243-250[本文引用:1]
LiuG, LuG, ZengL, Wang GL. Two broad-spectrum blast resistance genes, Pi9(t) and Pi2(t), are physically linked on rice chromosome 6. Mol Genet Genomics, 2002, 267: 472-480[本文引用:1]
[13]
朱小源, 杨健源, 刘景梅, 司徒志谋, 康金平, 胡学应, 朱敏记, 罗森辉, 杨祁云, 林佩珍, 曾列先, 姜先芽, 陈深. 广东水稻品种抗稻瘟病性分析与利用策略. 广东农业科学, 2006, (5): 34-37Zhu XY, Yang JY, Liu JM, Si-Tu Z M, Kang J P, Hu X Y, Zhu M J, Luo S H, Yang Q Y, Lin P Z, Zeng L X, Jiang X Y, Chen S. Evaluation on resistance of rice varieties in guangdong to rice blast and strategy for its utilization. Guangdong Agric Sci, 2006, (5): 34-37 (in Chinese with English abstract)[本文引用:1]
[14]
杨健源, 康金平, 黄显良, 姜先芽, 赖添奎, 陈深, 李传瑛, 曾列先, 朱小源. 稻瘟病田间自然诱发病圃的建立及其在抗性鉴定中的应用. 广东农业科学, 2007, (9): 59-61Yang JY, Kang JP, Huang XL, Jiang XY, Lai TK, ChenS, Li CY, Zeng LX, Zhu XY. Establishment of natural rice blast nursery and its application on the resistance evaluation of rice varieties. Guangdong Agric Sci, 2007, (9): 59-61 (in Chinese with English abstract)[本文引用:1]
[15]
吴双清, 王林, 卿明凤. 水稻区试品种抗稻瘟病鉴定结果分析. 中国稻米, 2011, 17: 53-55Wu SQ, WangL, Qing MF. Identification and analysis of regional tested rice varieties resistant to rice blast. China Rice, 2011, 17: 53-55 (in Chinese)[本文引用:1]
[16]
李求文, 杨隆维, 袁利群, 许敏. 持久抗稻瘟病杂交水稻新三系及组合选育与应用研究进展. 云南农业大学学报, 2006, 21: 276-282Li QW, Yang LW, Yuan LQ, XuM. Breeding and the applied research progress of hybrid Rice of new three line and combination of persistent resistance of blast. J Yunnan Agric Univ, 2006, 21: 276-282 (in Chinese with English abstract)[本文引用:2]
[17]
The 3000 rice genomes project. The 3000 rice genomes project. Gigascience, 2014, 3: 7[本文引用:1]
[18]
Li JY, WangJ, Zeigler RS. The 3000 rice genomes project: new opportunities and challenges for future rice research. Gigascience, 2014, 3: 8[本文引用:1]
[19]
湖北省质量技术监督局. 农作物品种区域试验抗病性鉴定操作规程(DB42/T 208-2001). 湖北省地方标准, 2001 年12月12日发布, 2002年1月1日实施Hubei ProvinceQuality and Technology SupervisionBureau. Resistance Identification Procedures of Crop Varieties for Regional Yield Test (DB42/T 208-2001). Hubei Local Stand ard, released on December 12, 2001 and effective on January 1, 2002 (in Chinese)[本文引用:1]
[20]
Bonman JM, Estrada BA, Band ong JM. Leaf and neck blast resistance in tropical lowland rice cultivars. Plant Dis, 1989, 73: 388-390[本文引用:1]
[21]
Zhuang JY, Ma WB, Wu JL, Chai RY, LuJ, Fan YY, Jin MZ, LeungH, Zheng KL. Mapping of leaf and neck blast resistance genes with resistance gene analog, RAPD and RFLP in rice. Euphytica, 2002, 128: 363-370[本文引用:1]
[22]
陈福如, 阮宏椿, 杨秀娟, 林时迟, 方琴, 严琰. 稻瘟病苗瘟叶瘟和穗颈瘟的相关性分析. 中国农学通报, 2006, 22: 440-443 (in Chinese with English abstract)Chen FR, Ruan HC, Yang XJ, Lin SC, FangQ, YanY. The correlation in seedlingblasts, leaf blasts and neck blasts of rice. Chin Agric Sci Bull, 2006, 22: 440-443[本文引用:1]
[23]
NoenplabA, VanavichitA, ToojindaT, SirithunyaP, TragoonrungS, SriprakhonS, VongsapromC. QTL mapping for leaf and neck blast resistance in Khao Dawk Mali10and Jao Hom Nin recombinant inbred lines. Scienceasia, 2006, 32: 133-142[本文引用:1]
[24]
MiahG, Rafii MY, Ismail MR, Puteh AB, Rahim HA, AsfalizaR, Latif MA. Blast resistance in rice: a review of conventional breeding to molecular approaches. Mol Biol Rep, 2012, 40: 2369-2388[本文引用:2]
[25]
李恩宇, 王悦, 陈光辉. 水稻稻瘟病抗性基因的定位及克隆研究进展. 作物研究, 2014, 28: 754-760Li EY, WangY, Chen GH. Advances on the mapping and cloning of blast resistance gene in rice. Crop Res, 2014, 28: 754-760 (in Chinese with English abstract)[本文引用:1]
[26]
KobayashiN, Telebanco-Yanoria M J, Tsunematsu H, Kato H, Imbe T, Fukuta Y. Development of new sets of international stand ard differential varieties for blast resistance in rice ( Oryza sativaL. ). Jpn Agric Res Q, 2007, 41: 31-37[本文引用:1]
[27]
何月秋, 唐文华. CO39 近等基因系抗稻瘟病性分析. 作物学报, 2001, 27: 838-841He YQ, Tang WH. Identification of CO39 near-isogenicl ines for rice blast resistance. Acta Agron Sin, 2001, 27: 838-841 (in Chinese with English abstract)[本文引用:]
[28]
Chou LS, LyonE, Wittwer CT. A comparison of high-resolution melting analysis with denaturing high-performance liquid chromatography for mutation scanning: cystic fibrosis transmembrane conductance regulator gene as a model. Am J Clin Pathol, 2005, 124: 330-338[本文引用:1]
[29]
KrypuyM, Newnham GM, Thomas DM, ConronM, DobrovicA. High resolution melting analysis for the rapid and sensitive detection of mutations in clinical samples: KRAS codon 12 and 13 mutations in non-small cell lung cancer. BMC Cancer, 2006, 6: 295[本文引用:1]
[30]
DoH, KrypuyM, Mitchell PL, Fox SB, DobrovicA. High resolution melting analysis for rapid and sensitive EGFR and KRAS mutation detection in formalin fixed paraffin embedded biopsies. BMC Cancer, 2008, 8: 142[本文引用:1]
[31]
Li JS, Wang XM, Dong RX, YangY, ZhouJ, YuC, ChengY, Yan CQ, Chen JP. Evaluation of high-resolution melting for gene mapping in rice. Plant Mol. Biol. Rep, 2011, 29: 979-985[本文引用:1]
[32]
罗文龙, 郭涛, 周丹华, 陈海英, 王慧, 陈志强, 刘永柱. 利用基于HRM的功能标记分析水稻 Wx和 fgr的基因型. 湖南农业大学学报, 2013, 39: 597-603Luo WL, GuoT, Zhou DH, Chen HY, WangH, Chen ZQ, Liu YZ. Analysis of rice genotypes rice Wx and fgr by HRM-based functional marker. J Hunan Agric Univ, 2013, 39: 597-603 (in Chinese with English abstract)[本文引用:1]
[33]
ShabanimofradM, Rafii MY, AshkaniS, Hanafi MM, Adam NA, Latif MA, Rahim HA, SahebiM. Analysis of SSR markers linked with brown planthopper resistance genes ( Bph) using high- resolution melting (HRM) in rice. Plant Omics, 2015, 8: 212-219[本文引用:1]
 , 陈凯
, 陈凯




{kind=link}
{kind=link}